
Submitted:
09 December 2025
Posted:
11 December 2025
You are already at the latest version
Abstract
Polygonatum odoratum is rich in polysaccharides, which are key bioactive components with significant pharmacological value. To optimize their extraction and utilization, it is crucial to identify the growth stage during which polysaccharides are most abundant. In this study, we investigated the seasonal dynamics of polysaccharide accumulation and protein expression in Polygonatum odoratum rhizomes via wide-targeted metabolomics and proteomics. The total polysaccharide content peaked in spring (13.3%), with the second highest level occurring in winter. A total of 610 differentially expressed proteins (DEPs) were identified, with the highest number of DEPs upregulated in spring. Weighted gene coexpression network analysis (WGCNA) revealed a module highly correlated with polysaccharide accumulation and enriched in pathways such as “starch and sucrose metabolism.” Key enzymes, including invertase (INV) and hexokinase (E2.7.1.4), were significantly upregulated in spring and positively correlated with polysaccharide biosynthesis. These results provide valuable insights into the optimal harvest time (spring) for Polygonatum odoratum and identify potential molecular targets for breeding high-polysaccharide varieties.
Keywords:
Polygonatum odoratum
; polysaccharides
; metabolomics
; proteomics
1. Introduction
Polygonatum odoratum is a perennial herb that is widely used in traditional Chinese medicine for its various health benefits, including immune regulation and anti-inflammatory and antioxidant properties [1,2]. The rhizomes of Polygonatum odoratum are highly valued as the primary medicinal component and contain various bioactive components, such as polysaccharides, steroidal glycosides, dipeptides, flavonoids, amino acids, and trace mineral elements. Among these, polysaccharides are considered one of the primary active components responsible for their medicinal effects [3]. Polysaccharides play crucial roles in plant growth, development, and stress responses, with their contents varying significantly depending on the plant’s growth stage and environmental conditions [4]. As important bioactive components in Polygonatum odoratum, polysaccharides exhibit immunomodulatory, antidiabetic, antiaging, antitumor, and antioxidant effects [5,6,7]. These pharmacological properties make them highly promising for applications in modern medicine and health supplements.
Polysaccharides are a class of complex carbohydrate compounds with high molecular weights formed by the dehydration of multiple monosaccharide molecules through condensation reactions. All carbohydrates and their derivatives that meet the definition of macromolecules can be classified as polysaccharides [8]. On the basis of the types of their constituent units, polysaccharides can be divided into two major categories: homopolysaccharides and heteropolysaccharides. Homopolysaccharides are composed of identical monosaccharide repeating units, such as starch and β-glucan; on the other hand, heteropolysaccharides are composed of two or more different monosaccharides, such as glucuronic acid and pectin. Polygonatum odoratum polysaccharides are a typical example of heteropolysaccharides and are primarily composed of various monosaccharides, including mannose, galactose, glucose, fructose, rhamnose, arabinose, and galacturonic acid [9].
Research has shown that the synthesis and accumulation of Polygonatum odoratum polysaccharides involve the action of multiple key enzymes, such as UDP-glucose pyrophosphorylase (UGPase), sucrose phosphorylase (SUS), and glucose transferases (GTs), which play crucial roles in the polysaccharide polymerization process [10]. Polysaccharide biosynthesis occurs through three main processes [5,11]. First, sucrose is converted into glucose-6-phosphate and fructose by β-fructofuranosidase, and fructose is subsequently converted into fructose-6-phosphate by hexokinase and fructokinase [12]. Glucose-6-phosphate isomerase catalyzes the isomerization of glucose-6-phosphate to glucose-1-phosphate, while UDP-glucose and GDP-mannose are produced from glucose-1-phosphate and fructose-6-phosphate precursors, respectively [12,13]. Second, several NDP-sugar interconversion enzymes catalyze the conversion of UDP-Glc or GDP-Man into other NDP sugars [14]. Finally, different glycosyltransferases remove monosaccharides from sugar nucleotides. The donor is bound to growing polysaccharide polymers, and these repeating units polymerize and output to form plant polysaccharides [15,16].
There are various methods for extracting Polygonatum odoratum polysaccharides, including hot water extraction, enzymatic extraction, and ultrasonic-assisted extraction. The traditional Polygonatum odoratum processing methods primarily include the “sugar-rubbing” method and steaming and roasting, both of which yield high contents of Polygonatum odoratum polysaccharides [17]. The modern processing method employed is ultrafine grinding, which significantly increases the polysaccharide content compared with traditional methods, providing new avenues for the extraction and application of Polygonatum odoratum polysaccharides. Subsequently, techniques such as ion exchange chromatography and gel permeation chromatography can be used to separate and purify Polygonatum odoratum polysaccharides further, yielding polysaccharide components with specific molecular weights and structures.
This study aimed to investigate the seasonal dynamic changes in polysaccharide content in Polygonatum odoratum rhizomes and explore the metabolic and proteomic changes associated with different growth stages. By integrating metabolomics and proteomics methods, we seek to comprehensively understand the changes in polysaccharide biosynthesis and accumulation [18,19]. This study also aimed to investigate the synthesis and regulatory mechanisms of secondary metabolites in Polygonatum odoratum at different growth stages, which may help optimize the harvest time to maximize the yield of polysaccharides and other major active components. To our knowledge, previous studies have rarely utilized multiomics integration methods to investigate changes in secondary metabolites in Polygonatum odoratum at different growth stages.
The results of this study lay the foundation for future genetic and metabolic engineering research on Polygonatum odoratum and other medicinal plants. By identifying key metabolic pathways and proteins involved in polysaccharide biosynthesis, this study opens new avenues for improving the quality and efficacy of Polygonatum odoratum as a medicinal resource.
2. Results
2.1. Total Polysaccharide Content in the Rhizomes of Polygonatum odoratum at Different Stages
The total polysaccharide content of Polygonatum root samples collected in different months is shown in Figure 1 and Table S1. The results revealed that the total polysaccharide content reached its peak (13.3%) in May (spring, SP). Analysis of the four seasons (February: winter, WI; May: spring, SP; August: summer, SU; November: fall, FA) revealed significant seasonal dynamic changes in the polysaccharide content of Polygonatum odoratum.
2.2. Polysaccharide Analysis of Polygonatum odoratum at Different Stages
Principal component analysis (PCA) was performed on the polysaccharides of Polygonatum odoratum at different growth stages, the results are shown in Figure 2A. The results revealed that principal component 1 (PC1) and principal component 2 (PC2) explained 47.14% and 22.29% of the variance, respectively [20]. Additionally, the figure shows that polysaccharides from Polygonatum odoratum samples collected in winter (WI) and autumn (FA) are highly similar, whereas those from spring (SP) and summer (SU) samples are significantly different. A heatmap based on polysaccharide-related metabolite data is shown in Figure 2B and Table S2. The heatmap further revealed differences in the polysaccharide metabolic profiles of Polygonatum odoratum at different growth stages [21]. These metabolites primarily include monosaccharides, disaccharides, trisaccharides, tetrasaccharides, monosaccharide phosphates, glycosamines, sugar alcohols, glycosides, sugar phosphates, sugar lactones, sugar acids, sugar lipids, and sugar esters. Heatmap analysis revealed that most polysaccharide-related metabolites reached their highest levels in spring (SP), whereas a few metabolites (such as certain disaccharides, glycosides, and sugar phosphates) presented relatively high levels in winter (WI).
2.3. WGCNA
Principal To investigate the associations between changes in Polygonatum odoratum polysaccharide content and gene expression, we analyzed proteomics data via weighted gene coexpression network analysis (WGCNA). By constructing a coexpression network, we identified gene modules significantly associated with Polygonatum odoratum polysaccharide content. The WGCNA trait heatmap in Figure 3 reveals correlations between different modules and polysaccharide content as well as other growth stage traits. Among these genes, the blue module was highly positively correlated with the polysaccharide content in rhizomes (correlation coefficient > 0.8), indicating that the DEPs within this module may play a key role in polysaccharide biosynthesis or regulation. These findings suggest that the blue module may influence the seasonal accumulation of Polygonatum odoratum polysaccharides by regulating carbohydrate metabolic pathways.
To analyze the internal structure of the blue module, we calculated the intramodule connectivity for each protein within the module. This metric measures the extent to which a protein interacts with other proteins within the module. Proteins with higher connectivity occupy more central positions in the network and are typically regarded as crucial hub proteins.Table S3 presents the top 20 core hub proteins identified based on connectivity. These hub proteins represent the core of the coexpression network, where their expression fluctuations decisively influence the network’s stability and functional output.
2.4. Identification and Annotation of DEPs
To better understand the mechanisms of polysaccharide synthesis and changes in Polygonatum odoratum, we performed proteomics measurements and analyses on rhizome samples of Polygonatum odoratum at different growth stages. We further analyzed the blue module via WGCNA and identified 610 Polygonatum odoratum polysaccharide-related proteins on the basis of a screening criterion of P < 0.05. To gain deeper insights into the functions of these differentially expressed proteins, we performed functional annotation using the KOG database and categorized the proteins into 23 groups as shown in Figure 4 [22,23].The most numerous DEPs were posttranslational modification, protein conversion, and chaperone proteins (67 DEPs), followed by general functional prediction (58 DEPs); signal transduction mechanisms (39 DEPs); energy production and conversion (38 DEPs); secondary metabolite biosynthesis, transport, and catabolism (37 DEPs); lipid transport and metabolism (31 DEPs); translation, ribosomal structure, and biosynthesis (30 DEPs); carbohydrate transport and metabolism (29 DEPs); and intracellular transport, secretion, and vesicle transport (21 DEPs).
2.5. Dynamic Changes in DEPs
Proteins showing a fold change of 1.5 or less were classified as differentially expressed proteins (DEPs), and those with a p value less than 0.05 were considered significantly different. After identification and screening, a total of 610 differentially expressed proteins (DEPs) were obtained from the 12 samples [24,25]. The number of DEPs obtained from stage-specific comparisons is shown in Figure 5. Specifically, in the SP_vs_WI comparison, 282 DEPs were identified, including 217 upregulated and 65 downregulated DEPs, suggesting that spring may activate many proteins related to growth and metabolism. In the SU_vs_WI comparison, a total of 215 DEPs were identified, with 167 upregulated and 48 downregulated DEPs, suggesting that metabolic activity remains relatively active in summer but is weaker than in spring. In the FA_vs_WI comparison, a total of 132 DEPs were identified, with 87 upregulated and 45 downregulated DEPs, reflecting a gradual slowing of metabolic activity in autumn. In the comparison between SU and SP, a total of 310 DEPs were identified, with 163 upregulated and 147 downregulated DEPs, indicating that some metabolic pathways may be inhibited in summer compared with spring. In the FA_vs_SP comparison, a total of 306 DEPs were identified, with 110 upregulated and 196 downregulated DEPs, suggesting that certain metabolic pathways may be inhibited in autumn compared with spring. In the FA_vs_SU comparison, a total of 109 DEPs were identified, with 23 upregulated and 86 downregulated and 86 downregulated , further supporting the trend of gradually decreasing metabolic activity with the progression of seasons. Compared with that in winter, the highest number of upregulated DEPs was observed in spring (217), which was significantly greater than that in other seasons. This finding is consistent with the results from metabolomics, which revealed that the polysaccharide content peaks in spring, suggesting that in spring, polysaccharide synthesis may be promoted by the activation of sugar metabolism-related proteins. The proportion of downregulated DEPs significantly increased from summer to autumn, implying that Polygonatum odoratum may adapt to environmental changes by regulating protein expression, reducing energy consumption, and preparing for winter dormancy.
2.6. Enrichment Analysis
To identify the KEGG pathways that were coenriched in the two omics studies, a coenriched analysis was performed on the polysaccharide metabolomics and proteomics data of Polygonatum odoratum, as shown in Figure 6A. The coenrichment bubble chart displays the KEGG pathways coenriched by the two omics methods, the number of differentially enriched metabolites and proteins in a specific pathway, the enrichment factor, and the significance level. By comprehensively analyzing the following bubble chart, it can be concluded that the polysaccharide metabolomics and proteomics of Polygonatum odoratum are enriched primarily in the starch and sucrose metabolic pathways.
GO is divided into three parts: molecular function, biological process, and cellular component. GO enrichment analysis, as shown in Figure 6B, indicates that polysaccharide-related differentially expressed proteins are primarily enriched in the molecular function categories of catalytic activity and binding function, the biological process categories of cellular processes and metabolic processes, and the cellular component category of cellular anatomical entities.
2.7. Analysis of Polysaccharide Biosynthesis Pathways
Through KEGG co-enrichment analysis, we integrated metabolomics and proteomics data to map polysaccharide biosynthesis pathways based on starch and sucrose metabolism pathways, as shown in Figure 7 [26]. During polysaccharide biosynthesis, multiple DEPs were identified, with INV and E2.7.1.4 being upregulated in spring. These proteins may contribute to polysaccharide accumulation. The relative abundance of DEPs exhibited seasonal differences, providing a foundation for understanding the seasonal variations in polysaccharides and other metabolites in Polygonatum odoratum.
2.8. Correlation Analysis Between DAMs and DEPs
To better understand the relationship between the metabolome and proteome of Polygonatum odoratum, we performed Pearson correlation analysis on the biosynthesis pathway of polysaccharides, analyzing 67 polysaccharide-related metabolites and 610 related DEPs. The correlation network diagram constructed by screening relevant coefficients (coefficient > 0.95) is shown in Figure 8. We found that a small number of DEPs were negatively correlated with polysaccharide accumulation, suggesting that these enzymes may play an inhibitory role in polysaccharide synthesis or participate in metabolic pathways opposite to polysaccharide accumulation; some DEPs were positively correlated with the accumulation of certain polysaccharides and negatively correlated with the accumulation of other polysaccharides, indicating that these DEPs may have complex regulatory mechanisms in the polysaccharide biosynthesis pathway, potentially participating in both promoting and inhibiting polysaccharide accumulation. The remaining DEPs were positively correlated with polysaccharide accumulation, indicating that these DEPs can promote the biosynthesis of Polygonatum odoratum polysaccharides. On the basis of the above correlation analysis results, we speculate that these DEPs may influence polysaccharide synthesis and accumulation by regulating the activity of key enzymes in the polysaccharide biosynthesis pathway.
3. Discussion
Polygonatum odoratum, a traditional Chinese herbal medicine, contains polysaccharides in its rhizomes, which are considered one of the primary active components. This study integrated metabolomics and proteomics methods to systematically analyze the seasonal variations in polysaccharide content and their molecular regulatory mechanisms in Polygonatum odoratum rhizomes across different growth seasons. The results not only revealed the key periods and regulatory networks for polysaccharide synthesis in Polygonatum odoratum but also provided an important theoretical basis for the quality control and cultivation of Polygonatum odoratum.
Through systematic seasonal sampling analysis, this study clarified the dynamic changes in polysaccharide content in the rhizomes of Polygonatum odoratum. The results revealed that the polysaccharide content of Polygonatum odoratum reached its peak (13.3%) in May (spring), which is consistent with the accumulation patterns of secondary metabolites in many medicinal plants. Spring is the rapid growth period for most plants, during which carbon metabolism is active, providing ample substrates for the synthesis of macromolecules such as polysaccharides. Our metabolomics data indicate that the contents of precursor substances such as monosaccharides and trisaccharides significantly increase at this time (Figure 2B), which is highly consistent with the increasing trend in polysaccharide content.
Notably, the polysaccharide compositions of the winter (WI) and autumn (FA) samples were highly similar (Figure 2A), which may be related to the relatively dormant state of the plants during these seasons. The polysaccharide content decreases in the summer (SU) samples, which is likely due to plants allocating more resources to growth metabolism rather than the synthesis of storage compounds under high-temperature conditions. This seasonal variation pattern suggests that seasonal factors should be particularly considered in the cultivation management and harvest timing selection of Polygonatum odoratum.
Through WGCNA, we identified a blue module significantly associated with polysaccharide content. The DEPs in this module may play a key role in polysaccharide biosynthesis or regulation, providing clues for understanding the molecular basis of polysaccharide biosynthesis in Polygonatum odoratum.
Among these DEPs, several key enzymes deserve special attention. The first is invertase (INV), which is significantly upregulated in spring (Figure 5). Invertase catalyzes the hydrolysis of sucrose into glucose and fructose, a step considered one of the rate-limiting steps in polysaccharide synthesis. Our results revealed that the expression level of INV was significantly positively correlated with polysaccharide content (Figure 8), strongly suggesting its key role in polysaccharide synthesis in Polygonatum odoratum. INV (invertase), as the key enzyme for sucrose hydrolysis, is upregulated in expression, which may be closely linked to the elongation of photoperiods and rising temperatures in spring. Research indicates that INV activity in various plants is regulated by light signaling pathways (such as those involving phytochrome-interacting factors (PIFs)) and temperature-sensitive transcription factors (such as HSFs). Higher average daily temperatures and abundant light in spring may activate these transcription factors, thereby promoting INV gene expression. This accelerates the conversion of sucrose into glucose and fructose, supplying ample precursors for polysaccharide synthesis. Additionally, increased soil moisture and nitrogen availability in spring may indirectly increase INV activity through hormonal signaling (e.g., gibberellins, auxins), further promoting the allocation of carbon sources toward stored polysaccharides. The second is hexokinase (E2.7.1.4), which promotes the production of UDP-glucose by phosphorylating glucose, and UDP-glucose is a direct precursor for the synthesis of various polysaccharides. Hexokinase (E2.7.1.4) is a key enzyme in hexose phosphorylation, and its springtime upregulation may reflect the high demand for energy and carbon skeletons during the rapid growth phase of Polygonatum odoratum. Like INV, hexokinase expression is also regulated by environmental factors. For example, in sugarcane and rice, hexokinase expression is significantly influenced by circadian rhythms and temperature fluctuations. The relatively moderate diurnal temperature variation in spring may favor maintaining stable hexokinase activity, thereby ensuring a continuous supply of nucleotide sugars such as UDP-glucose. Notably, the expression pattern of hexokinase in Polygonatum odoratum differs from that in some temperate medicinal plants, such as Panax ginseng: hexokinase expression peaks in autumn in ginseng, potentially because of its physiological strategy of accumulating storage compounds to withstand severe cold. In contrast, Polygonatum odoratum tends to rapidly synthesize polysaccharides in spring to support vegetative growth and reproductive preparation. This divergence may reflect species-specific ecological adaptation mechanisms.
Additionally, we identified several enzymes potentially involved in polysaccharide modification, such as glycosyltransferases and glycosidases. The expression of these enzymes also exhibited distinct seasonal variations (Figure 5), suggesting that they may play important roles in regulating the structural and functional diversity of polysaccharides. These findings provide new directions for understanding the biosynthetic pathways of Polygonatum odoratum polysaccharides.
By integrating metabolomics and proteomics data, we constructed a regulatory network for polysaccharide synthesis in Polygonatum odoratum (Figure 8). Correlation analysis revealed complex regulatory relationships between DEPs and polysaccharide metabolites (Figure 8). Most DEPs (e.g., INV and E2.7.1.4) were positively correlated with polysaccharide content, suggesting that these proteins may directly participate in the polysaccharide synthesis process. However, we also observed several interesting phenomena: some DEPs were negatively correlated with polysaccharide content, suggesting that these proteins may be involved in polysaccharide degradation or competitive metabolic pathways; others exhibited dual regulatory effects, implying the existence of more refined feedback regulatory mechanisms. In the network diagram, protein Cluster-15784.0.p1 stands out because of its strong positive correlation with the highest number of polysaccharide metabolites, including various monosaccharide phosphates and oligosaccharides. These findings suggest that this protein may serve as a pivotal hub in the polysaccharide synthesis network of Polygonatum odoratum. We hypothesize that it may function as a glycosyltransferase with multiple substrate specificities or as a signaling node regulating the activity of several downstream enzymes. This protein could synergistically promote the flow of multiple carbohydrate branch pathways, thereby efficiently directing precursor substances toward polysaccharide synthesis. Therefore, Cluster-15784.0.p1 is considered a highly promising molecular target. Future functional validation through overexpression experiments could identify this gene as a key factor in the breeding of Polygonatum odoratum varieties with high polysaccharide contents. Conversely, negatively correlated DEPs within the network also carry significant biological implications. These proteins, which are negatively correlated with polysaccharide accumulation, are likely not “detrimental factors” but rather manifestations of the fine-tuned metabolic balance regulation of the plant. These enzymes may initiate polysaccharide degradation—for example, certain glycosidases or amylase enzymes may activate once polysaccharide synthesis reaches a threshold, preventing excessive accumulation or recycling of carbon sources for other physiological activities. or participants in competing pathways. These DEPs may contribute to cell wall component synthesis (cellulose, lignin) or redirect carbon toward other secondary metabolites (flavonoids, saponins), thereby competing with polysaccharide synthesis for shared substrate pools. During spring, while polysaccharide synthesis dominates, mild activation of these competing pathways may aid in cell wall construction and structural maintenance. These genes may also function as regulators of energy and reducing power balance, with some negatively correlated DEPs potentially involved in respiration or oxidative stress responses. When carbohydrate metabolism becomes excessive, the upregulation of these proteins helps maintain intracellular ATP and NADPH homeostasis, preventing metabolic stress caused by accelerated carbon flux.
This complex regulatory network may reflect the adaptive strategies of plants to environmental changes. For example, under suitable growth conditions in spring, plants tend to accumulate stored polysaccharides; under high environmental stress, they may degrade polysaccharides to provide energy and carbon skeletons. This flexible metabolic regulatory capacity is highly important for the survival and accumulation of bioactive components in Polygonatum odoratum.
In terms of molecular mechanisms, the key enzymes identified in this study also have conserved functions in polysaccharide synthesis in other plants. However, certain DEPs unique to Polygonatum odoratum (such as some glycosyltransferase subtypes) may be associated with the unique structure of its polysaccharides. These findings provide new insights into the commonalities and characteristics of polysaccharides in different medicinal plants.
The results of this study have important practical implications. First, spring is the critical period for polysaccharide accumulation in Polygonatum odoratum, providing a scientific basis for determining the optimal harvest period. Second, the identified key DEPs can serve as molecular markers for the breeding and quality evaluation of superior Polygonatum odoratum varieties. Additionally, these findings provide potential targets for enhancing polysaccharide content in Polygonatum odoratum through genetic engineering.
Future research could validate the functions of key DEPs via gene editing technology; investigate the regulatory mechanisms of polysaccharide synthesis influenced by environmental factors (such as temperature, light, and moisture); conduct comparative studies of Polygonatum odoratum from different regions to analyze the impact of geographical variation on polysaccharide synthesis; and explore the relationship between polysaccharide structure and pharmacological activity.
4. Materials and Methods
4.1. Plant Materials
In a standardized experimental field in Nanling County, Wuhu city, Anhui Province (31°33′N, 118°38′E), the rhizomes of Polygonatum odoratum were planted for three years. Fresh Polygonatum odoratum rhizome samples were collected on February 2, 2022 (winter, abbreviated as WI), May 2 (spring, abbreviated as SP), August 2 (summer, abbreviated as SU), and November 2 (fall, abbreviated as FA). The samples were identified by Professor Fu Hongwei of Zhejiang Sci-tech University of Technology and stored at the College of Life Sciences and Medicine, Zhejiang Sci-tech University. The experimental field was managed with uniform fertilization, irrigation, and pesticide application measures. Polygonatum odoratum rhizomes were harvested in the third year after planting. At each sampling, 20 fresh rhizomes of Polygonatum odoratum were excavated from five fixed points (four corners and the central intersection point) in the experimental field and subjected to uniform processing. The rhizomes were washed with purified water, cut into 0.5 cm pieces, rapidly frozen in liquid nitrogen, and stored at -80 °C for subsequent experiments.
4.2. Broad-Target Metabolomics Analysis
Biological samples were first dried via a freeze dryer (Scientz-100F) and then ground into powder via an MM 400 grinder (Retsch, China) [27]. Fifty milligrams of powder (using an MS105DM electronic balance, Sartorius, Germany) was accurately weighed, and 1.2 mL of prechilled 70% methanol aqueous solution containing an internal standard was added to reach -20 °C. The sample was subjected to six vortex treatments followed by centrifugation, and the supernatant was collected and filtered for subsequent UPLC‒MS/MS analysis. Metabolomic analysis was performed by Met-ware Biotechnology Co., Ltd. (Wuhan, China). Metabolite peak area integration was conducted for all the samples via mass spectrometry, and the metabolite peaks were corrected. For group comparisons, differentially abundant metabolites (DAMs) were screened on the basis of P values (P < 0.05) and absolute Log2FC values (|Log2FC| ≥ 1.0). For multigroup comparisons, DAMs were identified on the basis of P values (P < 0.05, via analysis of variance). Additionally, metabolic pathways associated with DAMs were retrieved from the Kyoto Encyclopedia of Genes and Genomes (KEGG) database.
4.3. Determination of Total Polysaccharide Content
The phenol‒sulfuric acid method was used to determine the polysaccharide content in dried Polygonatum odoratum rhizome samples [28,29]. Each sample (February, May, August, and November) was mixed with 100 mL of distilled water, and the dried powder (0.2 g) was extracted twice at 100 °C in boiling distilled water for 1 hour each time. After extraction, the sample was precipitated by adding 95% ethanol (10 mL). The sample was then centrifuged, and the precipitate was dissolved in distilled water (50 mL) containing 4% phenol (1 mL) and sulfuric acid (7 mL). The absorbance of each sample was measured at 490 nm via a UV spectrophotometer (JASCO Corporation, Japan). The polysaccharide content was determined via three biological replicates.
4.4. Proteomic Analysis
Proteins were extracted from the samples via the acetone precipitation method. The specific steps were as follows: the samples were ground into a powder in liquid nitrogen and homogenized, after which an extraction mixture containing 100 mM Tris-HCl (pH 7.6), 1 mM phenylmethylsulfonyl fluoride (PMSF), and 2 mM ethylenediaminetetraacetic acid (EDTA) was added. Next, the protein sample was boiled for 15 minutes, sonicated in an ice bath for 10 minutes to lyse the cells, and centrifuged to obtain a clear protein mixture. Four times the volume of frozen acetone was added to the protein mixture, which was then incubated at -20 °C overnight to precipitate the proteins. The precipitate was then collected by centrifugation at 4 °C. The precipitate was washed with cold acetone and dissolved in 8 M urea. Finally, the protein concentration was determined via the BCA method according to the kit instructions.
For each sample, an equal amount of protein was used for trypsin digestion. 8 M urea was added to the supernatant to a volume of 200 µL, which was then treated with 10 mM DTT at 37 °C for 45 minutes for reduction, followed by treatment with 50 mM iodacetamide (IAM) at room temperature in the dark for 15 minutes for alkylation. Next, four volumes of ice-cold acetone were added, and the peptide segments were precipitated at -20 °C for 2 hours. After centrifugation, the protein precipitate was air-dried and resuspended in 200 µL of 25 mM ammonium bicarbonate solution. Three microliters of trypsin (trypsin-to-protein mass ratio of 1:50, Promega) was added, and the mixture was digested overnight at 37 °C. After digestion, the peptide segments were desalted via a C18 column (IonOpticks, Australia), dried in a vacuum concentrator, concentrated via vacuum centrifugation, and finally dissolved in 0.1% (v/v) formic acid solution.
4.5. Bioinformatics Analysis
WGCNA: WGCNA was performed on differentially expressed metabolites and proteins to identify coexpressed gene modules and explore the associations between gene networks and the phenotype of interest, as well as the core genes in the network [30]. WGCNA was conducted via MetWare Cloud (a free online data analysis platform (https://cloud.metware.cn/)).
KEGG pathway enrichment analysis: Bioinformatics analysis of differentially expressed metabolites and proteins on the basis of the KEGG biological pathway database to understand the biological pathways enriched by metabolites and proteins [27,31,32,33].
5. Conclusions
Through integrated metabolomics and proteomics analyses, this study systematically revealed the seasonal patterns and molecular regulatory mechanisms underlying polysaccharide accumulation in Polygonatum odoratum rhizomes. The results confirmed that spring (May) was the peak accumulation period (reaching 13.3% content), with a significant decline in summer, whereas autumn and winter presented similar polysaccharide compositions. Metabolomic data revealed significant accumulation of precursor substances such as monosaccharides and trisaccharides in spring, providing raw materials for polysaccharide synthesis. Proteomic analysis further revealed that key enzymes such as invertase (INV) and hexokinase (E2.7.1.4) were significantly upregulated in spring, and their expression levels were highly positively correlated with polysaccharide content.
WGCNA revealed a blue module highly positively correlated with polysaccharide content, where hub proteins (e.g., Cluster-15784.0.p1) may play central roles in the polysaccharide synthesis network. This module was significantly enriched in pathways related to “starch and sucrose metabolism.” This study also revealed that some DEPs were negatively correlated with polysaccharide accumulation, suggesting potential regulatory mechanisms involving metabolic balance and resource reallocation during seasonal growth.
These findings provide scientific support for the rationality of spring harvesting of Polygonatum odoratum and offer potential molecular targets for breeding high-polysaccharide varieties. Future research should further elucidate the regulatory network of polysaccharide synthesis in Polygonatum odoratum through gene functional validation and environmental regulation studies.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org.
Author Contributions
Conceptualization, Y.Z. and S.L.; methodology, Y.Z.; software, S.L. and D.F.; validation, Y.Z., S.L. and D.F.; formal analysis, Y.Z.; investigation, Y.Z. and S.L.; resources, H.F.; data curation, Y.Z. and S.L.; writing—original draft preparation, Y.Z.; writing—review and editing, H.F. and H.Z.; visualization, Y.Z.; supervision, H.F. and H.Z.; project administration, H.F. and H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yang, L. Y.; Yang, Q. W.; Zhang, L. P.; Ren, F. X.; Zhouyao Zhang, Z. Y.; Jia, Q. J. Integrated Metabolomics and Transcriptomics Analysis of Flavonoid Biosynthesis Pathway in Polygonatum cyrtonema Hua. Molecules 2024, 29, 2248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. W.; Li, X. Y.; Yu, D. H.; Yang, Z. Y.; Shen, Z. X.; Meng, Y. S.; Ding, Y. L.; Li, Y. Botany, chemistry, bio-activity, and application of Polygonatum odoratum (Mill.) Druce: a comprehensive review. Naunyn-Schmiedeberg’s Archives of Pharmacology. 2025. [Google Scholar] [CrossRef]
- Wang, C.; Yu, D. H.; Ruan, J. X.; Wang, X. F.; Liu, S. J. Research Progress on Chemical Components, Pharmacological Effects of Polygonatum odoratum and Prediction Analysis of Quality Markers. Chinese Journal of Modern Applied Pharmacy 2025, 42(1), 158–172. [Google Scholar] [CrossRef]
- Song, C. Research Progress on the Gene Regulatory Network of Plant Polysaccharide Biosynthesis. Botanical Research 2025, 14(3), 151–159. [Google Scholar] [CrossRef]
- Zhang, S. X.; Shi, Y. Y.; Huang, L. Q.; Wang, C. K.; Zhao, D. R.; Ma, K. L.; Wu, J. W.; Peng, D. Y. Comparative transcriptomic analysis of rhizomes, stems, and leaves of Polygonatum odoratum (Mill.) Druce reveals candidate genes associated with polysaccharide synthesis. Gene 2020, 744, 144626. [Google Scholar] [CrossRef]
- Wang, J.; Su, H. Y.; Han, H. P.; Wang, W. S.; Li, M. C.; Zhou, Y. B.; Li, Y.; Li, M. F. Transcriptomics Reveals Host-Dependent Differences of Polysaccharides Biosynthesis in Cynomorium songaricum. molecules 2022a, 27(1), 44. [Google Scholar] [CrossRef]
- Gong, D. Y.; Li, B.; Wu, B.; Fu, D. R.; Zesheng Li, Z. S.; Wei, H. B.; Guo, S. X.; Gang Ding, G.; Wang, B. C. The Integration of the Metabolome and Transcriptome for Dendrobium nobile Lindl. in Response to Methyl Jasmonate. Molecules 2023, 28, 7892. [Google Scholar] [CrossRef]
- Li, N.; Yu, X.; Yu, Q. H.; Wang, M. Research progress on stability of polysaccharides in traditional Chinese medicine. China journal of Chinese materia medica 2019, 44(22), 4793–4799. [Google Scholar] [CrossRef]
- Xu, S.; Bi, J. L.; Jin, W. F.; Fan, B. L.; Qian, C. Q. Determination of polysaccharides composition in Polygonatum sibiricum and Polygonatum odoratum by HPLC-FLD with pre-column derivatization. Heliyon 2022, 8, e09363. [Google Scholar] [CrossRef]
- Pan, G.; Jin, J.; Liu, H.; Zhong, C.; Xie, J.; Qin, Y. H.; Zhang, S. H. Integrative analysis of the transcriptome and metabolome provides insights into polysaccharide accumulation in Polygonatum odoratum (Mill.) Druce rhizome. PeerJ 2024, 12, e17699. [Google Scholar] [CrossRef]
- Wang, J.; Su, H. Y.; Wu, Z. B.; Wang, W. S.; Zhou, Y. B.; Li, M. F. Integrated Metabolites and Transcriptomics at Different Growth Stages Reveal Polysaccharide and Flavonoid Biosynthesis in Cynomorium songaricum. Int J Mol Sci 2022b, 23(18). [Google Scholar] [CrossRef]
- Richez, C.; Boetzel, J.; Floquet, N.; Koteshwar, K.; Stevens, J.; Badet, B.; Badet-Denisot, M-A. Expression and purification of active human internal His6-tagged L-glutamine: D-Fructose-6P amidotransferase I. Protein Expression & Purification 2007, 54, 45–53. [Google Scholar] [CrossRef]
- Decker, D.; Menga, M.; Gornickaa, A.; Hoferb, A.; Wilczynskab, M.; Kleczkowskia, L. A. Substrate kinetics and substrate effects on the quaternary structure of barley UDP-glucose pyrophosphorylase. Phytochemistry 2012, 79, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y. B.; Huang, J. L.; Gu, X. G.; Bar-Peled, M.; Xu, Y. Evolution of Plant Nucleotide-Sugar Interconversion Enzymes. PLOS one 2011, 6, e27995. [Google Scholar] [CrossRef] [PubMed]
- Lairson, L. L.; Henrissat, B.; Davies, G. J.; Withers, S.G. Glycosyltransferases: Structures, Functions, and Mechanisms. Annual Review of Biochemistry 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Chen, J. D.; Duan, X. J.; Li, Y. P.; Tao, Z. M. Comparative Transcriptome Profiling Reveals Two WRKY Transcription Factors Positively Regulating Polysaccharide Biosynthesis in Polygonatum cyrtonema. International Journal of Molecular Sciences 2023, 24(16), 12943. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, L.; Shan, X.; Zhao, T. T.; Cui, M. R.; Hao, W. Q.; Wei, B. B. Untargeted components and in vivo metabolites analyses of Polygonatum under different processing times. Lwt 2023, 173. [Google Scholar] [CrossRef]
- Yang, D. F; Du, X. H.; Yang, Z. Q.; Liang, Z. S.; Guo, Z. X.; Liu, Y. Transcriptomics, proteomics, and metabolomics to reveal mechanisms underlying plant secondary metabolism. Engineering in Life Sciences 2014, 14, 456–466. [Google Scholar] [CrossRef]
- Yang, B. X.; Li, Q.; Cheng, K. S.; Fang, J.; Mustafa, G.; Pan, J. J.; Xing, B. C.; Lv, Q. D.; Zhang, L.; Cheng, K. J. Proteomics and metabolomics reveal the mechanism underlying differential antioxidant activity among the organs of two base plants of Shiliang tea (Chimonanthus salicifolius and Chimonanthus zhejiangensis). Food Chemistry 2022, 385, 132698. [Google Scholar] [CrossRef]
- Fang, X. X.; Wang, H. Y.; Zhou, X. T.; Zhang, J.; Xiao, H. X. Transcriptome reveals insights into biosynthesis of ginseng polysaccharides. BMC Plant Biology 2022, 22, 594. [Google Scholar] [CrossRef]
- Xu, S. P.; Zhang, Y.; Liang, F.; Jiang, S. H.; Niu, S. Y.; Wang, X. M.; Zhou, Y. R.; Cui, B.; Yuan, X. Y. Metabolomic and transcriptomic analyses reveal the mechanism of polysaccharide and secondary metabolite biosynthesis in Bletilla striata tubers in response to shading. International Journal of Biological Macromolecules 2024, 279. [Google Scholar] [CrossRef]
- Wu, J.; Meng, X. X.; Jiang, W. M.; Wang, Z. J.; Zhang, J.; Meng, F.; Yao, X. Y.; Ye, M. J.; Yao, L.; Wang, L. H.; Yu, N. J.; Peng, D. Y.; Xing, S. H. Qualitative Proteome-Wide Analysis Reveals the Diverse Functions of Lysine Crotonylation in Dendrobium huoshanense. Frontiers in plant science 2022, 822374. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z. H.; Wu, M. M.; Yang, T. C.; Nan, X. Y.; Zhang, S. Y.; Zhang, L.; Jin, L. M. Integrated transcriptomic and proteomic analyses uncover the early response mechanisms of Catharanthus roseus under ultraviolet-B radiation. Journal of Photochemistry & Photobiology, B: Biology 2024, 252, 112862. [Google Scholar] [CrossRef]
- Zheng, X. H.; Xiao, H. B.; Su, J. Q.; Chen, D.; Chen, J. N.; Chen, B. H.; He, W. J.; Chen, Y. Q.; Zhu, J. M.; Fu, Y. J.; Ouyang, S. Y.; Xue, T. Insights into the evolution and hypoglycemic metabolite biosynthesis of autotetraploid Cyclocarya paliurus by combining genomic, transcriptomic and metabolomic analyses. Industrial Crops & Products 2021, 173, 114154. [Google Scholar] [CrossRef]
- Liu, D.; Dong, M. Y.; Li, M. F.; Jin, L.; Wei, J. H.; Paré, P.W. Metabolite and transcriptomic changes reveal the ripening process in Sinopodophyllum hexandrum fruit. Industrial Crops & Products 2023, 206. [Google Scholar] [CrossRef]
- Ning, L. Y.; Xu, Y. S.; Luo, L.; Gong, L. M.; Liu, Y. M.; Wang, Z.; Wang, W. Integrative analyses of metabolome and transcriptome reveal the dynamic accumulation and regulatory network in rhizomes and fruits of Polygonatum cyrtonema. BMC Genomics 2024, 25(1), 706. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X. R.; Zhang, X. Z.; Duan, S. D.; Su, X.; Fan, Z. H.; Hao, L. H.; Xiang, D. Y.; Chen, D. F.; Niu, S. C. Integrated metabolomic and transcriptomic analysis reveals the mechanism of high polysaccharide content in tetraploid Dendrobium catenatum Lindl. Industrial Crops & Products 2024, 212, 118391. [Google Scholar] [CrossRef]
- Yue, F. F.; Zhang, J. R.; Xu, J. X.; Niu, T. F.; Lü, X.; Liu, M. S. Effects of monosaccharide composition on quantitative analysis of total sugar content by phenol-sulfuric acid method. Frontiers in Nutrion 2022, 9, 963318. [Google Scholar] [CrossRef]
- Zhang, Y. W.; Shi, Y. C.; Zhang, S. B. Metabolic and transcriptomic analyses elucidate a novel insight into the network for biosynthesis of carbohydrate and secondary metabolites in the stems of a medicinal orchid Dendrobium nobile. Plant Diversity 2023, 45(3), 326–336. [Google Scholar] [CrossRef]
- Zheng, G. Q.; Wang, Z. Q.; Wei, J. R.; Zhao, J. H.; Zhang, C.; Mi, J. J.; Zong, Y.; Liu, G. H.; Wang, Y.; Xu, X.; Zeng, S. H. Fruit development and ripening orchestrating the biosynthesis and regulation of Lycium barbarum polysaccharides in goji berry. Int J Biol Macromol 2024, 254 Pt 2, 127970. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, M. M.; Zhao, L. T.; Chen, L.; Ding, Z. Y. Novel Insights into the Mechanism Underlying High Polysaccharide Yield in Submerged Culture of Ganoderma lucidum Revealed by Transcriptome and Proteome Analyses. Microorganisms 2023, 11(3). [Google Scholar] [CrossRef]
- Zhu, J.; Cai, Y. M.; Yang, L. Y.; Li, X.; Luo, S. F.; Zeng, X. H.; Zhang, Y. C.; Huang, W. C. Differential metabolic analysis of Bletilla striata and its mutants based on widely targeted metabolomics and transcriptomics. Industrial Crops & Products 2023, 204. [Google Scholar] [CrossRef]
- You, H. Q.; Li, S. Q.; Chen, Y. W.; Lin, J. J.; Wang, Z. X.; Dennis, M.; Li, C. Y.; Yang, D. F. Global proteome and lysine succinylation analyses provide insights into the secondary metabolism in Salvia miltiorrhiza. Journal of Proteomics 2023, 288, 104959. [Google Scholar] [CrossRef]
- Ma, R. X.; Zhang, M.; Yang, X. H.; Guo, J.; Fan, Y. L. Transcriptome analysis reveals genes related to the synthesis and metabolism of cell wall polysaccharides in goji berry (Lycium barbarum L.) from various regions. Journal of the Science of Food and Agriculture 2023, 103, 7050–7060. [Google Scholar] [CrossRef]
- Phu, L. P.; Thi, T. C. L.; Thi, T. H. V.; Thanh, T. N.; Zhang, Z. S.; Zeng, R. Z.; Xie, L.; Minh, N. N.; Vuong, T. H. T.; Tran, D. X.; Tran, D. K. Comparative Transcriptome Analysis and Expression of Genes Associated with Polysaccharide Biosynthesis in Dendrobium officinale Diploid and Tetraploid Plants. Agronomy 2024, 14(1), 69. [Google Scholar] [CrossRef]
- Yang, B. X.; Zhong, Z. H.; Wang, T. T.; Ou, Y. T.; Tian, J. K.; Komatsuc, S.; Zhang, L. Integrative omics of Lonicera japonica Thunb. Flower development unravels molecular changes regulating secondary metabolites. Journal of Proteomics 2019, 208, 103470. [Google Scholar] [CrossRef]
- Ma, Y. P.; Devi, M.J.; Song, L. H.; Gao, H. D.; Jin, L.; Cao, B. Multi-omics integration analysis reveals metabolic regulation of carbohydrate and secondary metabolites during goji berry (Lycium barbarum L.) maturation. Postharvest Biology and Technology 2024, 218. [Google Scholar] [CrossRef]
Figure 1.
Polysaccharide content of the cultivated variety Polygonatum odoratum.
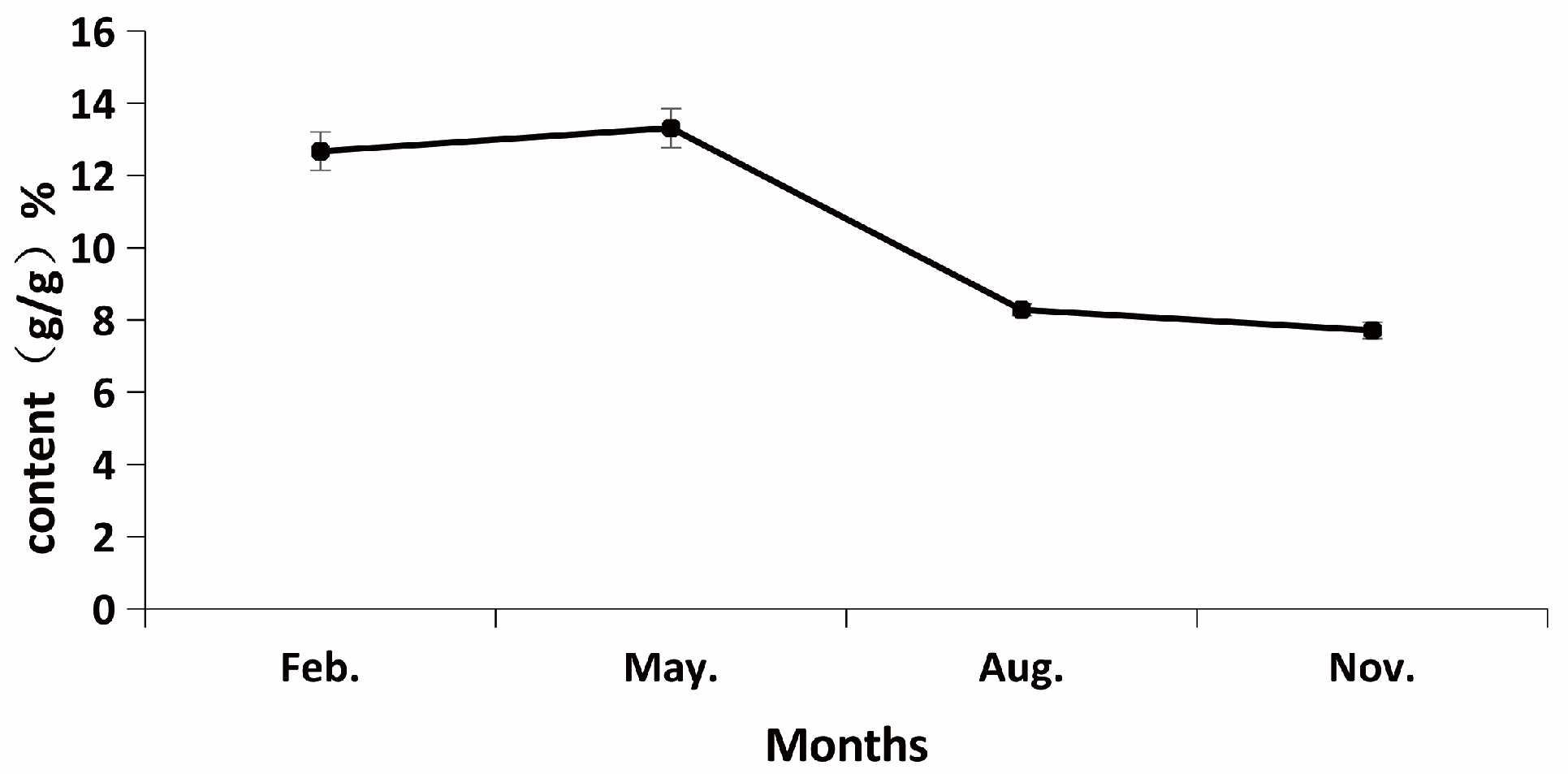
Figure 2.
A: Principal component analysis of polysaccharides from Polygonatum odoratum at different growth stages; B: Heat map of Polygonatum odoratum polysaccharides at different growth stages.
Figure 2.
A: Principal component analysis of polysaccharides from Polygonatum odoratum at different growth stages; B: Heat map of Polygonatum odoratum polysaccharides at different growth stages.
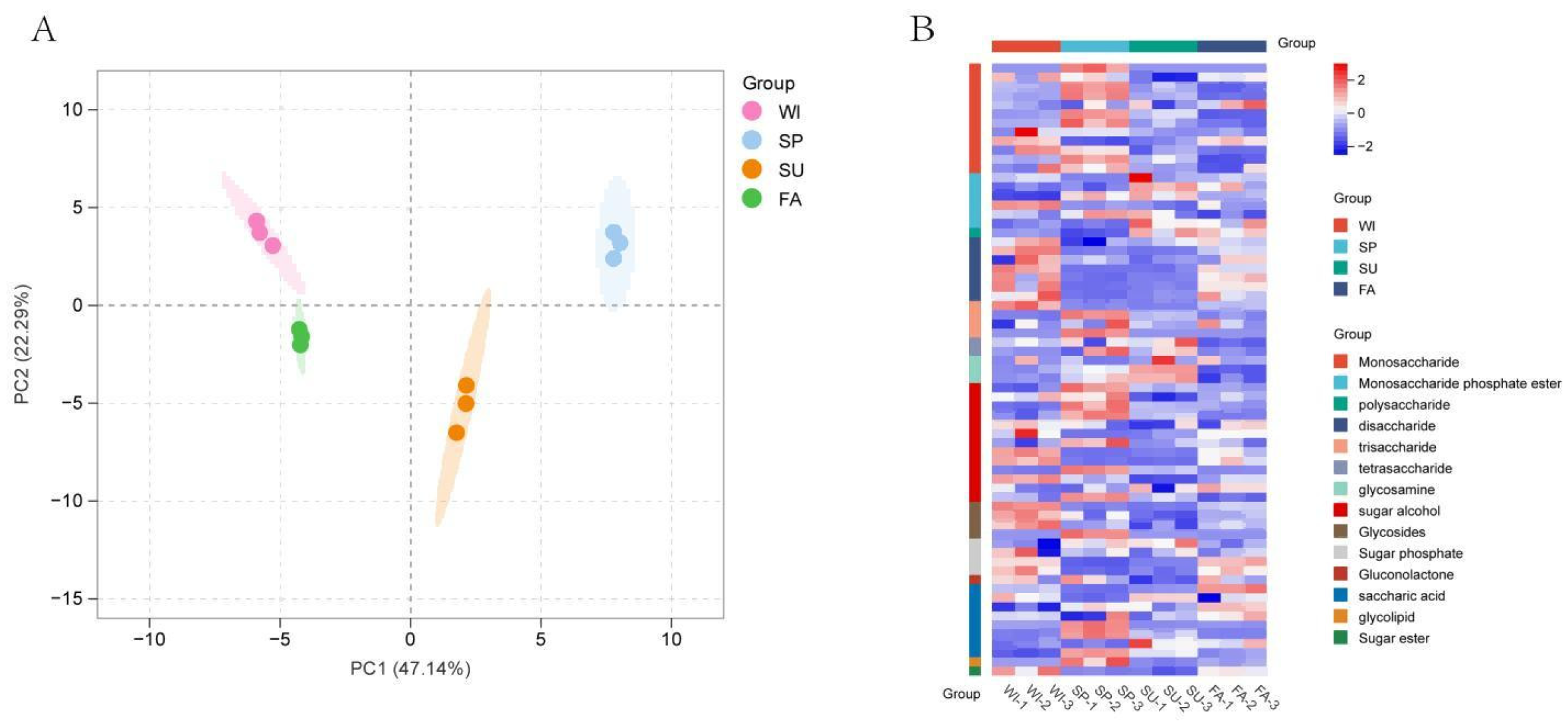
Figure 3.
Heat map of traits analyzed by WGCNA.
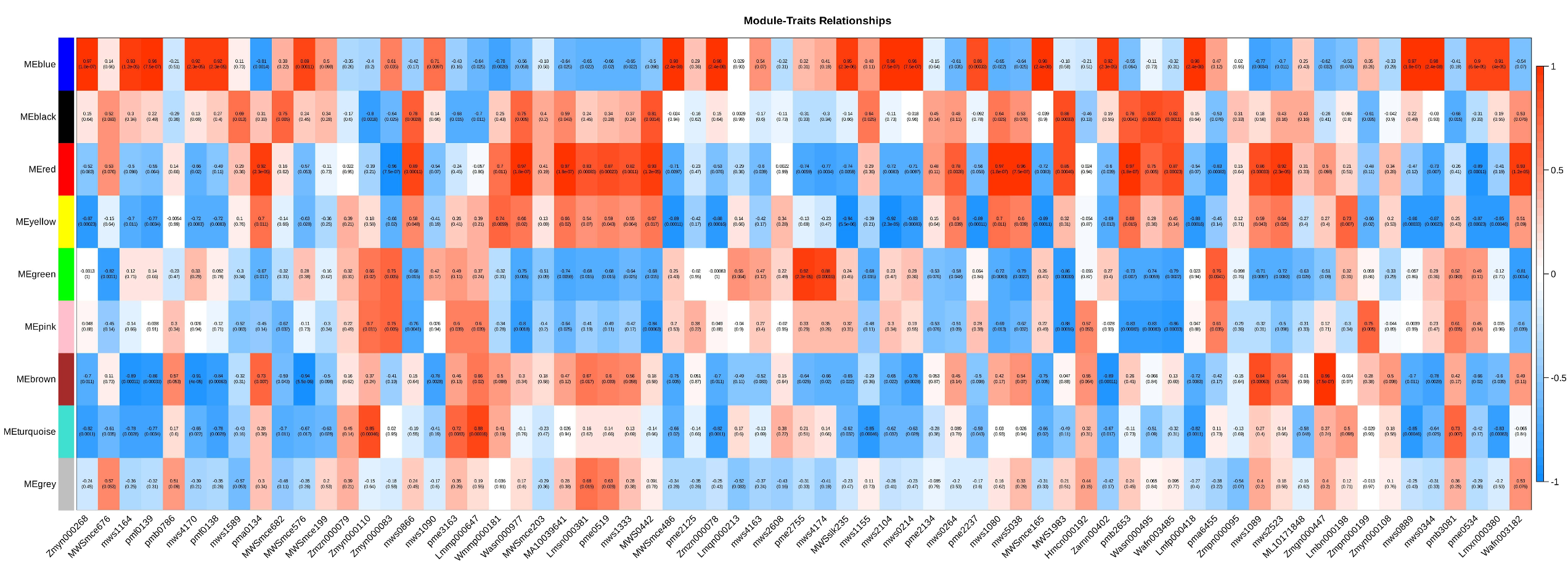
Figure 4.
KOG functional annotation of DEPs.
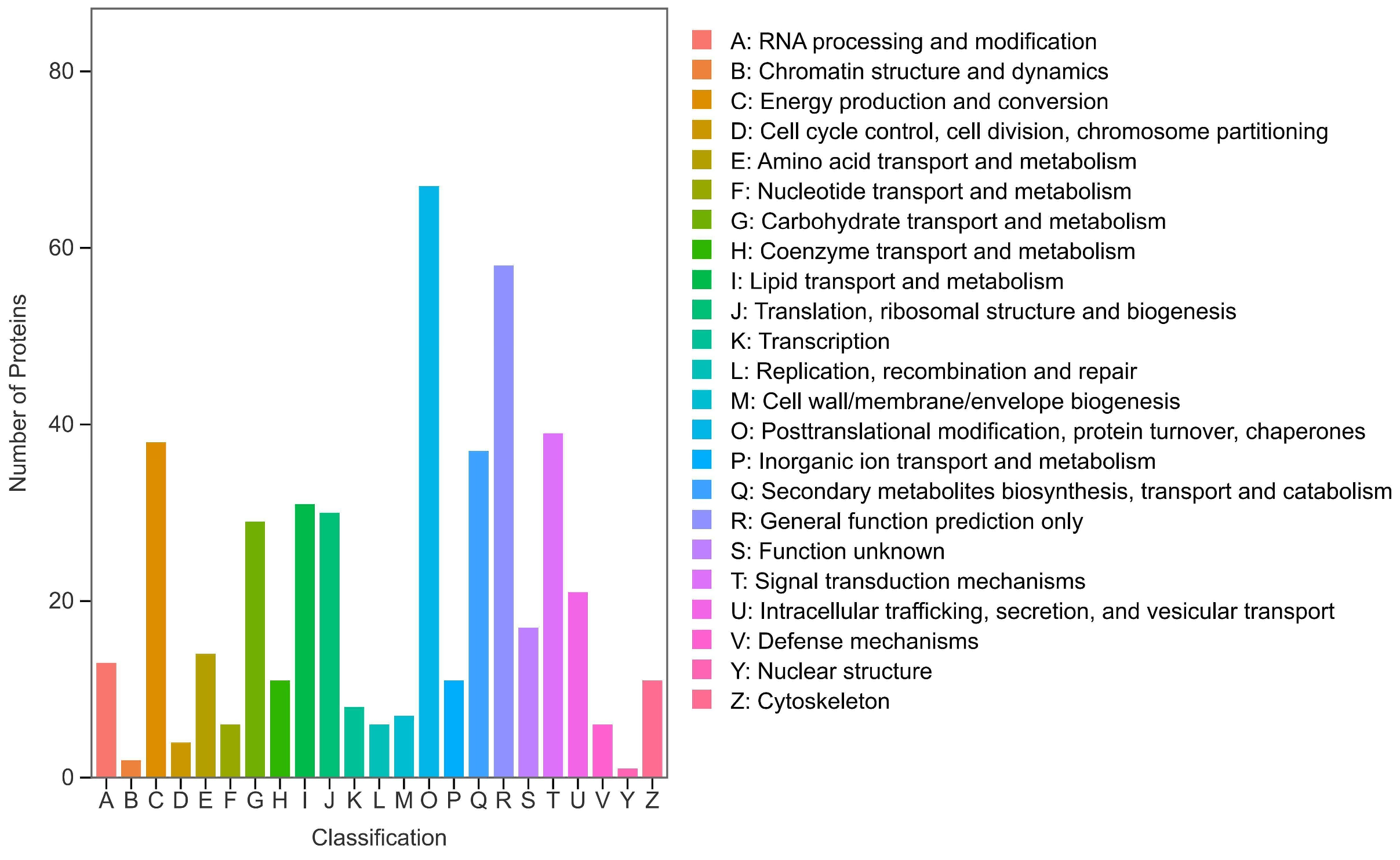
Figure 5.
Number of DEPs at different growth stages.
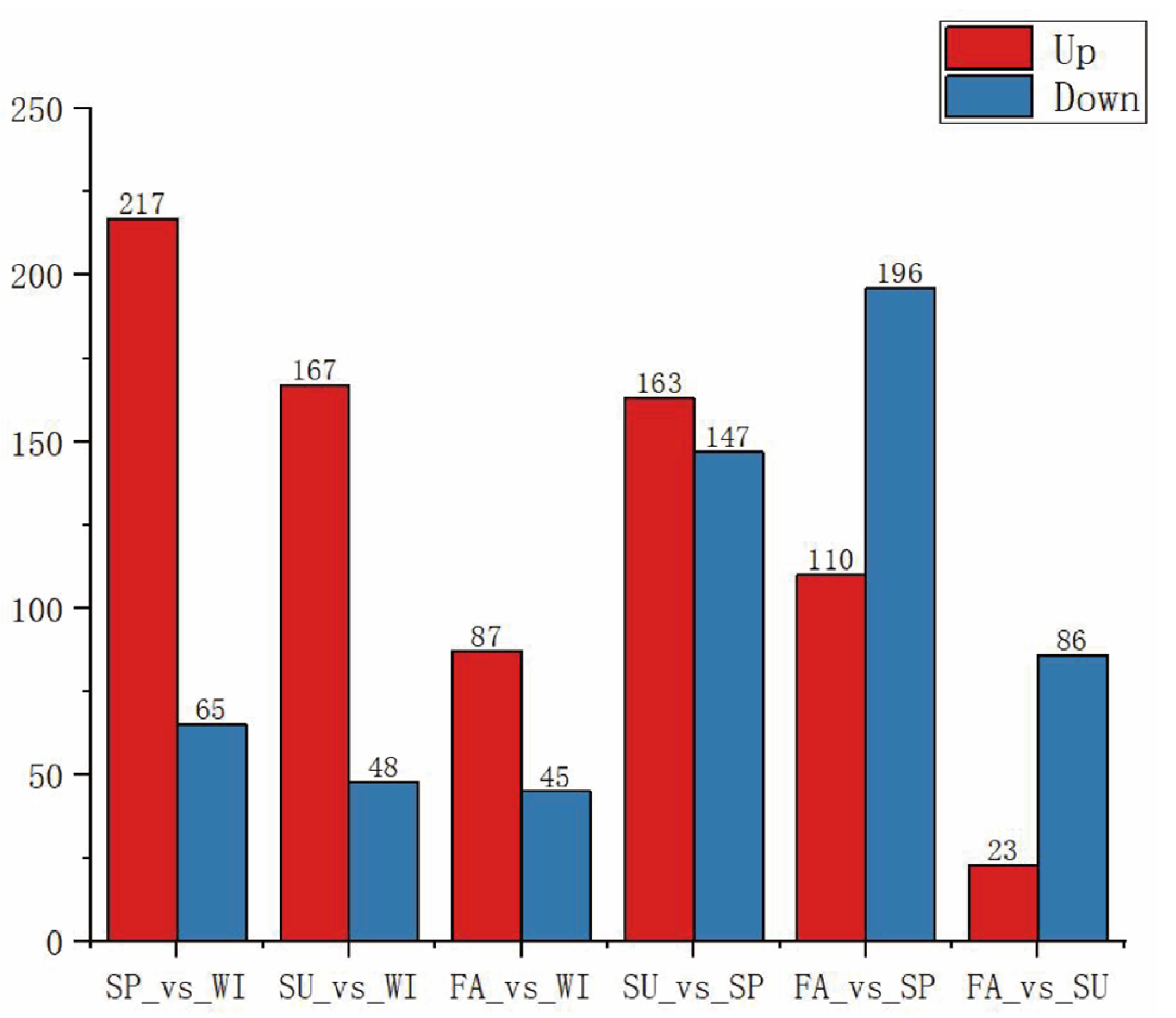
Figure 6.
A: Co-enrichment bubble chart of polysaccharide metabolomics and proteomics; B: GO enrichment analysis of DEPs.
Figure 6.
A: Co-enrichment bubble chart of polysaccharide metabolomics and proteomics; B: GO enrichment analysis of DEPs.
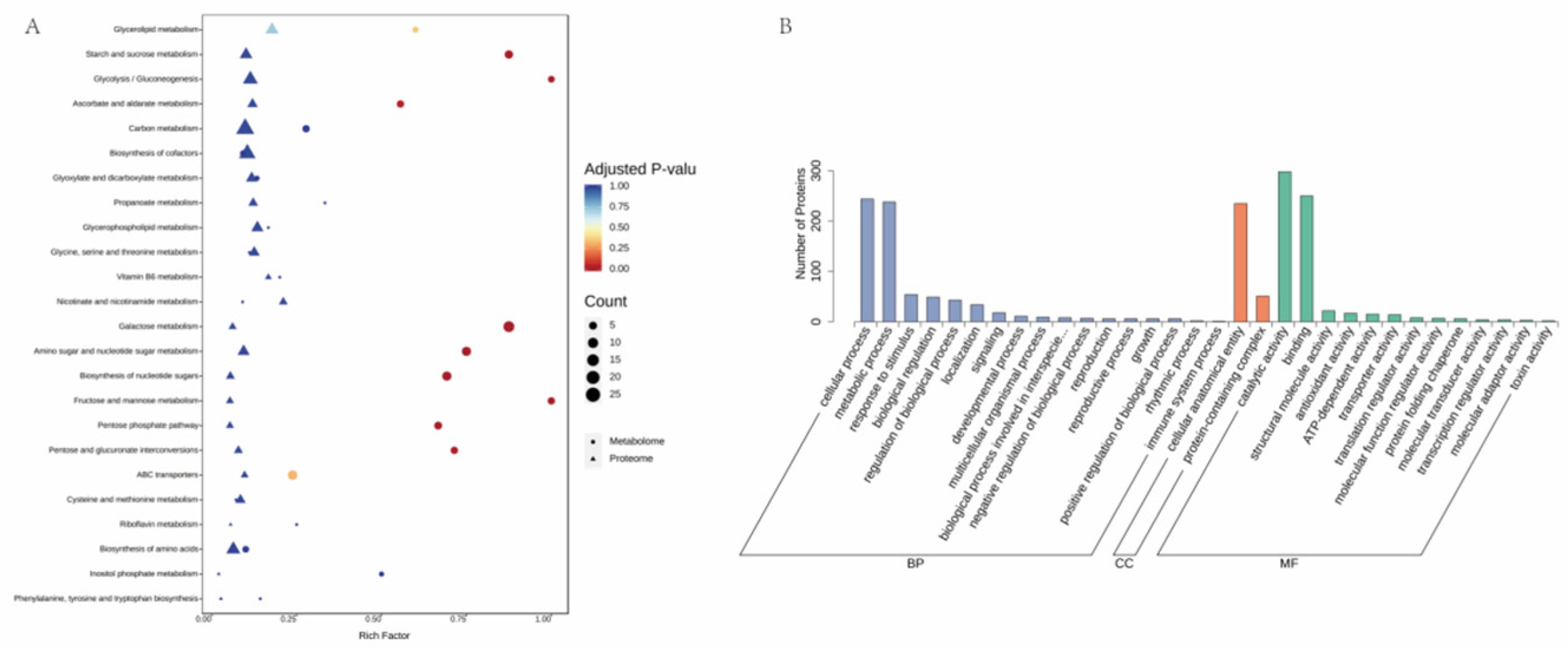
Figure 7.
Polysaccharide biosynthesis pathway diagram.
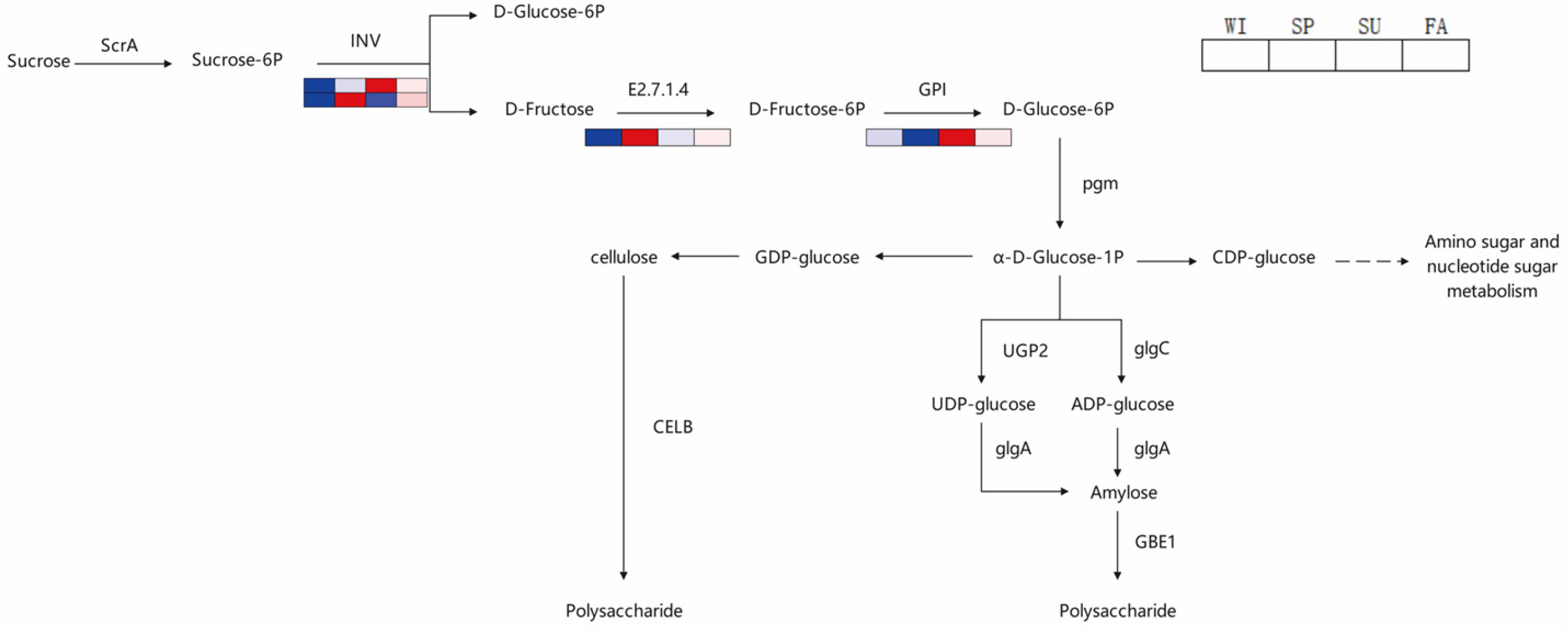
Figure 8.
Pearson correlation analysis between polysaccharide-related metabolites and DEPs.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



