
Submitted:
03 November 2025
Posted:
04 November 2025
You are already at the latest version
Abstract
Sustainable extraction methods represent a key strategy in green chemistry and nutraceutical development, aiming to replace conventional solvent-based techniques while maintaining extract quality and safety. This study compared pistachio ( Pistacia vera L.) extracts obtained by ultrasound-assisted extraction (UAE) and a traditional solvent-based protocol, focusing on compositional features and biological effects. Ex-tracts were characterized for their chemical profiles, and their impact on HCT-116 co-lon-derived cells was evaluated through viability assays and gene expression analysis. The UAE-derived extract, richer in carbohydrates, promoted higher cell proliferation after 72 h, whereas the classical extract upregulated HMOX-1, suggesting activation of antioxidant defense pathways. Moreover, UAE treatment downregulated GLUT2 ex-pression while increasing IL-6 and IL-10 transcripts, indicating a possible carbohy-drate-driven immunometabolic response. Overall, these findings highlight both the advantages and limitations of green extraction approaches: while environmentally sustainable and efficient, ultrasound-assisted protocols may modify extract composi-tion in ways that influence biological responses. Optimization of extraction parameters is therefore essential to ensure a balance between ecological sustainability, composi-tional integrity, and biological safety.

Keywords:
Pistacia vera
; colorectal cancer
; green extraction methods
; glucose metabolism
; inflammatory pathway
; HCT‐116 cells
1. Introduction
In recent decades, there has been growing interest in the use of plant-based compounds for health promotion, particularly those derived from edible sources with recognized nutritional value. Among these, pistachio (Pistacia vera L.) has attracted considerable attention not only for its organoleptic and dietary qualities, but also for its promising nutraceutical potential. Economically, pistachio production represents a significant agricultural sector, generating both edible nuts and substantial quantities of lignocellulosic by-products such as hulls, leaves, and shells, which are now being explored for their biotechnological valorization[1]. Biologically, Pistacia vera is a dioecious tree of the Anacardiaceae family and is the only species of the Pistacia genus to produce edible nuts. Its seeds are a rich source of various phytochemicals, including phenolic acids, flavonoids (e.g., catechins, anthocyanins, flavonoids), stilbenes, terpenoids, and antioxidant vitamins such as tocopherols. These bioactive constituents contribute to a wide range of health benefits, including antioxidant, anti-inflammatory, antimicrobial, vasoprotective, and cardioprotective effects[2]. These properties have led to their recommended inclusion in cardioprotective diets such as the Mediterranean diet and to evidence-based correlations with improvements in lipid profiles, glycemic control, and oxidative stress biomarkers in both healthy and at-risk populations. Consequently, pistachio extracts have been increasingly integrated into functional foods and nutraceutical formulations[3]. In parallel with this trend, the methods used to obtain bioactive compounds from natural matrices have been the subject of analysis. Traditional extraction techniques, such as maceration, the Soxhlet method, or reflux extraction, often involve large volumes of organic solvents and high energy consumption, raising environmental and safety concerns. Conversely, the development of green extraction technologies, including ultrasound-assisted extraction (UAE), microwave-assisted extraction (MAE), and supercritical fluid extraction (SFE), offers a more sustainable approach. These methods aim to reduce the use of organic solvents, extraction times, and energy requirements, while maintaining or even improving the yield and quality of bioactive compounds[4,5]. Green extraction methods are gaining popularity for the isolation of plant-derived nutraceuticals due to their environmental benefits. However, co-extraction of non-target compounds can compromise the desired bioactivity, particularly in pathological contexts such as cancer. Several plant-derived preparations have already been investigated in oncology: for instance, Vinca alkaloids[6] (vincristine, vinblastine) Catharanthus roseus[7] and Camptothecin derivatives [8]are widely used as chemotherapeutics. In addition, nutraceutical-grade plant extracts such as curcumin[9] (from Curcuma longa) and resveratrol[10] (from grapes and berries) are under clinical evaluation for their adjuvant roles in cancer therapy. In this context, the inadvertent presence of pro-inflammatory or glucose-like molecules in poorly characterized extracts could alter their biological activity, potentially shifting their effect from protective to pro-tumorigenic. This underscores the need for rigorous compositional screening during extract development, especially for applications in oncology. The nutraceutical market has expanded significantly over the past 30–40 years, driven by consumer demand for natural, health-promoting products. However, this rapid growth is increasingly clashing with emerging environmental standards and policies, particularly those outlined in the European Green Deal, which promote sustainable production and reduced environmental impact across all industrial sectors. The need to reconcile commercial growth with ecological responsibility has intensified the search for environmentally friendly alternatives for the extraction of bioactive compounds[11]. In this context, this study aims to investigate the influence of extraction methodology on the chemical composition and biological activity of pistachio-derived extracts. By comparing traditional and eco-sustainable extraction techniques, the study aims to determine how methodological differences impact antioxidant activity and biomolecular responses. Ultimately, this work contributes to the ongoing effort to develop sustainable, effective, and safe strategies for producing high-value nutraceuticals, particularly for applications in complex disease contexts such as cancer. This study investigates how green extraction methods can affect extract composition, thus altering nutraceutical efficacy—originally attributed to the presence of monounsaturated fatty acids (MUFA) and polyunsaturated fatty acids (PUFA) in Pistacia vera extracts—and instead promote inflammatory responses. Overall, the data suggest that, while green extraction offers clear environmental benefits, insufficient selectivity can compromise nutraceutical precision.
2. Materials and Methods
2.1. Pistachio Preparation and Extraction
Pistachio samples were obtained from two distinct geographical origins: Spain and Bronte (Sicily, Italy). Prior to polyphenolic extraction, all samples underwent a standardized defatting procedure to remove lipophilic components that could interfere with subsequent extraction and analysis. Following established protocols [12]finely chopped pistachio samples (5 g) were subjected to lipid extraction using n-hexane (10 mL) under continuous agitation for 24 hours at room temperature. The mixture was subsequently allowed to settle by gravity separation for 2-3 hours, after which the supernatant containing dissolved lipids was carefully decanted and discarded. The remaining defatted pistachio material was filtered under vacuum using a Buchner funnel equipped with filter paper to remove residual n-hexane and achieve complete solvent elimination. This defatting step served as a prerequisite for both extraction methodologies employed in this study, ensuring that subsequent polyphenolic extraction procedures would target the desired hydrophilic and amphiphilic bioactive compounds without interference from the substantial lipid fraction naturally present in pistachio kernels.
First Method (Green Method). According to the literature [13] the two different pistachio samples previously defatted (1 g each, weighed on an analytical digital balance), originating from Spain and Italy, were extracted using a 45% v/v ethanol/water solution by means of Ultrasound-Assisted Extraction (UAE). The extraction was carried out in an ultrasonic thermostated bath at 50 °C for 15 minutes. After extraction, the samples were centrifuged at 5000 rpm for 30 minutes at 5 °C to remove residual fats and coarse particulates. The recovered supernatant was then concentrated using a rotary evaporator, and the resulting fraction was stored in a freezer. Ultimately, 88 mg of extract were obtained from the Bronte pistachio sample, and 115 mg from the Spanish one.
Second Method. According to the literature [12]the two previously defatted pistachio samples (5 g each, accurately weighed on an analytical digital balance) from Spain and Italy were mixed with 50 mL of methanol containing 0.1% (v/v) HCl. The mixtures were sonicated for 15 minutes and subsequently centrifuged (5000 rpm, 10 min, 4 °C). The resulting pellets were extracted twice more under the same conditions. The methanolic fractions were pooled and concentrated using a rotary evaporator. The residues were then dissolved in 20 mL of distilled water and extracted four times with 20 mL of ethyl acetate in a separating funnel. The combined organic phases were dried over Na₂SO₄ for 20 minutes. Ultimately, 90 mg of extract were obtained from the Bronte pistachio sample and 163 mg from the Spanish one.
At the end of the extraction process, in both cases, the final product was resuspended with methanol.
2.2. Cell Culture and Treatments
A human colorectal cancer cell line (HCT-116, Cat. No. CCL-247, ATCC®, Manassas, VA, USA) was cultured in McCoy’s 5A (Cat. No. MS029E1001, Biowest, Nuaillé, France) medium supplemented with 10% of fetal bovine serum (FBS, Cat. No. S1560-500, Biowest, Nuaillé, France) and penicillin/streptomycin (50 μg/ml, Cat. No. L0022-100, Biowest, Nuaillé, France) at 37°C in a controlled atmosphere with 5% CO2 according to the supplier's instructions. The lyophilized pistachio extracts, obtained using a green extraction method using a conventional extraction method, were resuspended in a buffer solution of 70% ethanol and phosphate- buffered saline (PBS, Cat. No. 10010023, Gibco™, Thermo Fisher Scientific, Waltham, MA, USA) in a 1:1 ratio to achieve a final stock concentration of 5 µg/µL. The day after seeding, cells were treated with pistachio extracts diluted in McCoy’s medium to a final concentration of 250 ng/µL.
2.3. Impact of Extracts on Cell Viability and Proliferation
Both pistachio extracts were solubilized in a 1:1 (v/v) mixture of PBS and 70% ethanol, and tested for their metabolic impact at a final concentration of 250 ng/μL[14,15]. Treatments were performed by adding 10 μL of extract solution directly into 96-well plates containing the cells.
HCT-116 cells were seeded at a density of 5,000 cells per well in McCoy’s 5A medium supplemented with 10% FBS, 1% Penicillin-Streptomycin, according to ATCC recommendations. Cells were treated with the green extract or the classic extract for 24, 48, and 72 hours. Untreated cells were included in each assay and used as controls.
Cell viability was assessed using the MTT assay[16]. At each time point, 10 μL of MTT solution (5 mg/mL in 1× PBS), prepared from powder (Cat. No. M5655, Sigma-Aldrich, St. Louis, MO, USA), was added to each well and incubated for 3 hours at 37 °C in a humidified incubator with 5% CO₂. Formazan crystals were solubilized with dimethyl sulfoxide (DMSO, Sigma-Aldrich), and absorbance was measured at 570 nm using a BioTek Synergy H1 Multimode Reader (Agilent Technologies, Santa Clara, CA, USA).
All experimental conditions and time points were performed in quadruplicate (technical replicates) and repeated in at least three independent biological experiments.
2.4. Impact on Inflammatory and Metabolic Pathways
HCT-116 cells were seeded in 6-well plates at a density of 200,000 cells per well and incubated overnight under standard culture conditions. The following day, cells were treated for 24 hours with either the green extract or the classic extract at a final concentration of 250 ng/μL. In addition to the two pistachio-derived extracts, a heteroarylethylene compound named PB4, previously synthesized and characterized in our laboratory[16,17,18], was included for comparative purposes. PB4 has been reported in our earlier publications to possess significant cytotoxic activity (IC₅₀= 0.32 μM [16]) and to markedly influence cellular oxidative and inflammatory processes[19]. In this work, it was tested at a concentration equal to half its IC₅₀ for 24 hours to simulate an inflammatory but non-lethal environment for the cells. After treatment, cell culture supernatants were removed, and cells were lysed directly in the wells by adding RLT buffer supplemented with β-mercaptoethanol, following the manufacturer’s instructions for the QIAGEN RNeasy Mini Kit (Ref. 74104, QIAGEN, 40724 Hilden, Germany). The lysates were processed immediately for total RNA extraction to allow subsequent analysis of genes involved in inflammatory and metabolic pathways.
2.5. RNA Extraction and cDNA Synthesis
RNA extraction was carried out following the manufacturer’s instructions provided by QIAGEN RNeasy Mini Kit (Ref. 74104, QIAGEN, 40724 Hilden, Germany). RNA was quantified using both the Eppendorf BioPhotometer® D30 and the fluorimeter Qubit RNA HS Assay Kit to evaluate purity and quantity of the initial sample, respectively (Ref. 32850, Invitrogen, 92008 Carlsbad, CA, USA). The samples with an A260/A280 ratio between 1.8 and 2.1 were considered suitable for downstream applications. For cDNA synthesis, 1 μg of total RNA was reverse transcribed using the QuantiTect Reverse Transcription Kit (Qiagen, Cat. No. 205311). This kit includes an integrated genomic DNA wipeout step to eliminate potential DNA contamination and a mix of oligo-dT and random primers to ensure efficient and unbiased cDNA synthesis. The resulting cDNA was stored at –20 °C.
2.6. RT-qPCR
Gene expression measurements were performed by means of mRNA quantification using a LightCycler® 480 II instrument (Roche, Switzerland) using QuantiTect SYBR Green PCR Kit ( Cat. No. 204143 QIAGEN, Germany) and cDNA per well (50 ng total per well). Real-time cycler conditions: Initial Activation Step, 15 minutes, 95 °C. 3-step cycling ×40 cycles: Denaturation, 15 seconds, 94 °C; Annealing, 30 seconds, 55 °C; Extension, 30 seconds, 72 °C (Fluorescence Acquisition in this step). As a negative control, a reaction in the absence of cDNA (no template control, NTC) was performed. All primers in this studio are QuantiNova LNA PCR Assay (Table 1, Cat.No. 249990, QIAGEN GmbH, Gilden, Germany). Target gene expression levels (GLUT2, IL-6, IL-10, HMOX-1) were normalized to the housekeeping gene RPLP0, and relative quantification was performed using the ΔΔCt method. Data analysis was carried out using the LightCycler® 480 software (version 1.5). The relative RNA expression level for each sample was calculated using the well-known 2−∆∆CT method in which the threshold cycle (CT) value of the gene of interest is compared to the CT value of our selected internal control (RPLP0)[20]. qRT-PCR amplifications were performed in triplicate. [21]
3. Results
3.1. Comparative Chemical Results Between the Extracts
Given that both extraction protocols utilize solvent systems with well-established affinity for bioactive polyphenolic compounds, we focused our comparative analysis on extraction efficiency as the primary discriminating factor for preliminary method assessment. The extraction yield for the first method (green method) is about 8.8% for Bronte and about 11.5% for Spain. The extraction yield for the second method (Classic Method) is about 1,8% for Bronte and about 3,26% for Spain. Based on the extraction yields, the ultrasound-assisted extraction (UAE) with ethanol/water (45% v/v) proved to be more efficient for both pistachio samples. In contrast, the second approach, consisting of an initial extraction with acidified methanol followed by liquid–liquid partitioning with ethyl acetate, was more complex and time-consuming. Although this method introduces additional purification steps (centrifugation, ethyl acetate partitioning, and drying over sodium sulfate), which may enhance selectivity by removing unwanted components and improving the purity of the final product, it resulted in lower overall yields.
3.2. Metabolic Impact of green and classic extracts
To evaluate the metabolic impact of both pistachio extracts on HCT-116 cells (Figure 1, Table S1), an MTT assay was conducted after 24, 48, and 72 hours of treatment at a final concentration of 250 ng/μL. At 24 hours, no significant difference in cell viability was observed between the green extract-treated and control (untreated) cells (average growth 104.8%, p-value > 0.05). After 48 hours, the green extract induced a modest but statistically significant reduction in cell viability, with treated cells displaying approximately 96% viability compared to control (p = 0.0141). At 72 hours, a marked decrease in viability was detected, with treated cells showing only 62.5% viability relative to control (p < 0.0001). However, this result was not associated with cytotoxicity. Bright-field microscopy revealed extensive cellular overgrowth and detachment in treated wells at 72 hours, suggesting that the reduced MTT signal was due to hyperproliferation-induced confluence and subsequent detachment, rather than direct cytotoxic effects of the extract.
As for the classic extract, after 24 hours treated cells exhibited a slight but statistically significant increase in metabolic activity compared to untreated controls, reaching 109.65% viability (p = 0.0226). At 48 hours, a significant reduction in cell viability was observed, with treated cells showing 82.13% viability relative to control (p < 0.0001). However, at 72 hours, cell viability returned to levels comparable to untreated cells (104.69%), with no statistically significant difference (p > 0.05). These findings indicate a transient inhibitory effect at 48 hours, followed by a recovery of metabolic activity over time.
3.3. Impact on Inflammation Pathway
The expression levels of HMOX-1, IL-6, and IL-10 were assessed by RT-qPCR in HCT-116 cells treated for 24 hours with the green extract or the classic extract at a final concentration of 250 ng/μL. Gene expression was normalized to the housekeeping gene RPLP0 and reported as fold change relative to untreated control cells (Figure 2, Table S2). Regarding cytokines, the green extract treatment induced a no significant increase in IL-6 expression (1.06) and a significant increase in IL-10 expression (5.04) relative to control. the classic extract, on the other side, significantly downregulates the expression of IL-6 (0.19) or IL-10 (0.27). These results suggest that the green and classic extracts exert distinct effects on gene expression, with the classic extract preferentially inducing an antioxidant response (HMOX-1), while the green extract promotes immunomodulatory cytokine expression (IL-6, IL-10). Treatment with the classic extract resulted in a marked upregulation of HMOX-1 expression, with a relative fold change of 4.85 compared to control. In contrast, HMOX-1 expression in the green extract-treated cells was 2.90.
3.4. Impact on Glucose Pathway
A marked modulation of glucose transporter expression was observed upon treatment with the green extract (Figure 3, Table S2). Specifically, GLUT2 transcript levels were drastically reduced (fold change: 0.15) compared to untreated controls, indicating a strong transcriptional downregulation. By contrast, cells treated with the classic extract maintained GLUT2 expression at levels comparable to control. This sharp decrease in GLUT2 expression with the green extract aligns with previous findings from our laboratory[21], where HCT-116 cells exposed to excess glucose or carbohydrate-rich substrates exhibited significant GLUT2 downregulation. The present data therefore reinforce the association between elevated extracellular carbohydrate availability and reduced GLUT2 transcription in this cancer cell model, suggesting that the green extract contains a higher fraction of glucose or carbohydrate-derived compounds than its classic counterpart.
4. Discussion
In recent years, increasing attention has been given to the development of sustainable extraction technologies. Within this framework, the European Green Deal initiatives have received considerable attention[22], particularly for promoting chemical practices that support environmental protection and sustainability [23]. Much attention was given to the promotion of sustainable technologies, including green extraction methods to reduce environmental impact.
The transition from conventional extraction methods using organic solvents (e.g., n-hexane or acidified methanol) to greener alternatives (e.g., ethanol, ultrasound, microwaves, or supercritical fluids) aligns closely with the principles of circular bioeconomy and environmental sustainability [24]. These eco-friendly techniques reduce toxic solvent usage and enhance the recovery of agro-industrial residues [25]. Nevertheless, it remains essential to assess extract selectivity and purity alongside environmental and economic benefits.
The differences in yields observed between classical and green extraction methods reflect fundamentally distinct extraction philosophies rather than mere variations in efficiency. Ultrasound-assisted extraction (UAE) employs a polar solvent system designed to recover a broad spectrum of water-soluble and moderate polar compounds, including sugars, organic acids, amino acids, and phenolic compounds[26]. This approach emphasizes maximal recovery of the total polar fraction from the plant matrix. In contrast, classical extraction incorporates sequential purification steps for selective compound isolation. Initial acidified methanol extraction targets phenolic compounds under conditions that enhance their solubility, followed by liquid-liquid partitioning with ethyl acetate to achieve selective separation[27]. Additional steps such as centrifugation, phase separation, and anhydrous drying systematically remove co-extracted polar interferents that could interfere with downstream applications[28].
Therefore, yield differences should be interpreted in terms of extraction selectivity rather than just efficiency. The higher recoveries observed with UAE likely reflect the co-extraction of non-target polar metabolites[29], whereas the lower yields from classical methods suggest enhanced selectivity for phenolic compounds, potentially resulting in extracts with higher specific bioactivity per unit mass. The UAE method, being faster, simpler, and aligned with green chemistry principles, represents a practical compromise between yield, operational simplicity, and environmental sustainability[30].
In our study, we analyzed pistachio extracts obtained using both green and classical extraction methods. Green extracts exhibited remarkably a probably high carbohydrate content, highlighting the differential selectivity of aqueous and ethanol-based extraction techniques. To investigate the biological consequences of these compositional differences, HCT-116 colon cancer cells were treated with the respective extracts. The green extract induced hyperproliferation after 72 hours, leading to rapid confluence and subsequent detachment, probably due to the high sugar content. This finding emphasizes potential risks in pathological contexts, where increased glucose availability may alter cellular metabolism and influence disease progression. Notably, the polyphenol-rich classical extract induced higher HMOX-1 expression[31] compared to both the sugar-rich green extract and the control. HMOX-1, an inducible enzyme regulated by the Keap1/Nrf2/ARE signaling pathway, responds to oxidative and electrophilic stress[32]. Paradoxically, polyphenols also trigger this endogenous antioxidant response, acting as both free radical scavengers and stimulators of cellular defense mechanisms [33,34,35]. Thus, HCT-116 cells treated with the classical extract exhibited strong activation of the Keap1/Nrf2/ARE pathway, reflecting a robust internal antioxidant response mediated by polyphenols.
Gene expression analysis revealed a pronounced modulation of glucose transport pathways in response to the different pistachio extracts. Notably, cells treated with the green extract exhibited a drastic downregulation of GLUT2 (fold change: 0.15), a key facilitative glucose transporter involved in maintaining intracellular glucose homeostasis[36]. Such a marked reduction suggests an adaptive cellular response aimed at limiting glucose uptake under conditions of high extracellular carbohydrate availability—consistent with the elevated sugar content previously identified in the green extract. This finding is particularly relevant in light of previous work from our research group, in which HCT-116 cells subjected to glucose deprivation markedly overexpressed GLUT2[37], enabling the selective internalization of 5-fluorouracil-functionalized nanoparticles, while cells exposed to a high-glucose medium showed a notable reduction in the expression of GLUT2, how it’s happened with green and classical extract (Figure 3). The current results therefore reinforce the notion that GLUT2 expression in HCT-116 cells is highly plastic and responsive to extracellular glucose levels, with potential implications for nutrient sensing and targeted drug delivery strategies. In parallel, the green extract treatment also led to increased expression of IL-6 and IL-10, cytokines associated with pro-inflammatory and immunomodulatory responses, respectively. The concurrent downregulation of GLUT2 and upregulation of these cytokines suggests a coordinated shift in immunometabolic programming, possibly driven by carbohydrate-induced metabolic stress[38]. This pattern was not observed with the classic extract, which maintained GLUT2 expression at control levels and displayed a distinct gene expression signature dominated by HMOX-1 induction.
In summary, while the increased carbohydrate content may provide energy substrates in non-pathological contexts, in cancer models such enrichment raises concerns due to the enhanced glycolytic metabolism of tumour cells. The increased availability of sugars associated with bioactive antioxidants could help athletes maintain oxidative metabolism and maintain redox balance during intense physical exertion, which is a nutraceutical benefit rather than a limitation[39,40,41]. However, the intended therapeutic or nutraceutical applications of pistachio extracts need to be carefully considered. The biological effects observed with the green extract may reflect differences in carbohydrate content, highlighting the importance of optimizing extraction selectivity. While a high carbohydrate content is beneficial as it provides a readily available source of energy, critical considerations need to be made for therapeutic applications, particularly in metabolic disorders or cancer patients, as the well-documented increased glycolytic metabolism of cancer cells (Warburg effect) [42] could inadvertently promote tumor growth.
While green extraction methods are certainly in line with sustainability goals, their potential to extract unwanted components requires further optimization[43,44]. Future research should focus on refining extraction parameters to maximize the selectivity of bioactive compounds while minimizing unwanted carbohydrate fractions. The use of advanced analytical techniques, such as metabolomics, could further elucidate the metabolic pathways affected by these extracts and help in the evaluation of their clinical and nutraceutical potential.
In summary, the strategic use and careful refinement of green extraction methods are crucial to achieving a balance between environmental sustainability, purity of extracts and therapeutic efficacy. This balance is crucial for the effective and safe use of pistachio-derived nutraceuticals in various clinical and nutritional contexts.
5. Conclusions
This study highlights the significant impact of extraction methodology on the chemical composition and biological activity of pistachio-derived extracts. Although green extraction methods are environmentally advantageous, this study revealed that they lead to high carbohydrate content, which induces hyperproliferative and pro-inflammatory responses in HCT-116 colon cancer cells. These effects were evidenced by increased cell confluence and detachment, downregulation of GLUT2, and upregulation of pro-inflammatory cytokines such as IL-6 and IL-10. In contrast, extracts obtained through classical methods demonstrated a stronger antioxidant profile, as reflected by enhanced HMOX-1 expression, likely mediated by polyphenol-induced activation of the Keap1/Nrf2/ARE pathway [45]. Collectively, these findings highlight the need for precise control over extraction parameters to ensure the safety and efficacy of pistachio-based nutraceuticals, particularly in pathological contexts such as cancer. In future applications, the therapeutic potential of these extracts in vivo models should be explored so that their biological activity can be validated, and systemic responses can be assessed. Additionally, industrial applications could benefit from optimized green extraction processes that preserve bioactivity while aligning with the sustainability goals of the Green Deal. Ongoing experiments will aim to complete cytokine profiling to further elucidate the immunomodulatory effects of the extracts. Moreover, advanced metabolomic approaches will be employed to better characterize the biochemical pathways affected by extract composition, ultimately guiding the development of safer and more targeted nutraceutical formulations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, TableS1; TableS2.
Author Contributions
Conceptualization, N.M.; methodology, G.Z., M.T., V.G., and G.C.; software, N.M.; validation, V.G. and N.M.; formal analysis, N.M.; investigation, G.Z., P.G.B., and M.T.; resources, C.B. and C.G.F.; data curation, G.Z., P.G.B., and M.T.; writing—original draft preparation, G.Z., P.G.B., M.T., and G.C.; writing—review and editing, P.G.B. and C.B.; visualization, C.B.; supervision, V.G., C.B., and N.M.; project administration, C.G.F. and N.M.; funding acquisition, C.G.F. and N.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants funded by Progetto PRIN 2022, avviso di riferimento D.D. 104 del 02/02/2022, piano di riferimento PNRR, Missione 4, Componente 2, Investimento 1.1, Project Title: FRASTUCA−CUP: E53D2301006 0006–Code: 2022WYFST2, UPB: 51723152021.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors would like to thank the BRIT laboratory at the University of Catania (Italy) for the valuable technical assistance and use of their laboratories, and Dr. Gaetano Savoca for his contribute.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| UAE | Ultrasound-Assisted Extraction |
| GC | Gas Chromatography |
| HPLC | High-Performance Liquid Chromatography |
| qPCR | Quantitative Polymerase Chain Reaction |
| RNA | Ribonucleic Acid |
| DNA | Deoxyribonucleic Acid |
| IL-6 | Interleukin 6 |
| IL-10 | Interleukin 10 |
| HMOX-1 | Heme Oxygenase 1 |
| GLUT2 | Glucose Transporter Type 2 |
| ROS | Reactive Oxygen Species |
| DMEM | Dulbecco’s Modified Eagle Medium |
| FBS | Fetal Bovine Serum |
| PBS | Phosphate Buffered Saline |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| SD | Standard Deviation |
| ANOVA | Analysis of Variance |
| CV | Coefficient of Variation |
References
- Elakremi, M.; Sillero, L.; Ayed, L.; ben Mosbah, M.; Labidi, J.; ben Salem, R.; Moussaoui, Y. Pistacia Vera L. Leaves as a Renewable Source of Bioactive Compounds via Microwave Assisted Extraction. Sustainable Chemistry and Pharmacy 2022, 29, 100815. [Google Scholar] [CrossRef]
- Ballistreri, G.; Arena, E.; Fallico, B. Influence of Ripeness and Drying Process on the Polyphenols and Tocopherols of Pistacia Vera L. Molecules 2009, 14, 4358–4369. Molecules 2009, 14, 4358–4369. [Google Scholar] [CrossRef]
- Grace, M.H.; Esposito, D.; Timmers, M.A.; Xiong, J.; Yousef, G.; Komarnytsky, S.; Lila, M.A. In Vitro Lipolytic, Antioxidant and Anti-Inflammatory Activities of Roasted Pistachio Kernel and Skin Constituents. Food Funct 2016, 7, 4285–4298. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, E.; Rezaei, A.; Piravivanak, Z.; Mirani Nezhad, S.; Rashidi Nodeh, H.; Safavi, M. Extraction of Phytosterols from the Green Hull of Pistacia Vera L. Var and Optimization of Extraction Methods. Journal of Medicinal plants and By-products 2025, 14, 252–259. [Google Scholar] [CrossRef]
- Bodoira, R.; Velez, A.; Rovetto, L.; Ribotta, P.; Maestri, D.; Martínez, M. Subcritical Fluid Extraction of Antioxidant Phenolic Compounds from Pistachio (Pistacia Vera L.) Nuts: Experiments, Modeling, and Optimization. J Food Sci 2019, 84, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Beljanski, M.; Beljanski, M.S. Selective Inhibition of in Vitro Synthesis of Cancer DNA by Alkaloids of Beta-Carboline Class. Exp Cell Biol 1982, 50, 79–87. [Google Scholar] [CrossRef]
- Saman, S.S.; Hanif, A.; Al-Rawi, S.S.; Ibrahim, A.H.; Iqbal, M.A.; Khalid, S.; Majeed, A.; Mahmood, A.; Ahmad, F. Catharanthus Roseus (L.) G. Don: A Herb with the Potential for Curing Breast Cancer. Next Research 2025, 2, 100218. [Google Scholar] [CrossRef]
- Venditto, V.J.; Simanek, E.E. Cancer Therapies Utilizing the Camptothecins: A Review of in Vivo Literature. Mol Pharm 2010, 7, 307–349. [Google Scholar] [CrossRef]
- Zoi, V.; Galani, V.; Lianos, G.D.; Voulgaris, S.; Kyritsis, A.P.; Alexiou, G.A. The Role of Curcumin in Cancer Treatment. Biomedicines 2021, 9, 1086. [Google Scholar] [CrossRef]
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int J Mol Sci 2017, 18, 2589. [Google Scholar] [CrossRef]
- Cheriyan, B.V.; Karunakar, K.K.; Anandakumar, R.; Murugathirumal, A.; kumar, A.S. Eco-Friendly Extraction Technologies: A Comprehensive Review of Modern Green Analytical Methods. Sustainable Chemistry for Climate Action 2025, 6, 100054. [Google Scholar] [CrossRef]
- Mandalari, G.; Tomaino, A.; Arcoraci, T.; Martorana, M.; Turco, V.L.; Cacciola, F.; Rich, G.T.; Bisignano, C.; Saija, A.; Dugo, P.; et al. Characterization of Polyphenols, Lipids and Dietary Fibre from Almond Skins (Amygdalus Communis L.). Journal of Food Composition and Analysis 2010, 23, 166–174. [Google Scholar] [CrossRef]
- Detti, C.; dos Santos Nascimento, L.B.; Brunetti, C.; Ferrini, F.; Gori, A. Optimization of a Green Ultrasound-Assisted Extraction of Different Polyphenols from Pistacia Lentiscus L. Leaves Using a Response Surface Methodology. Plants 2020, 9, 1482. [Google Scholar] [CrossRef] [PubMed]
- Seifaddinipour, M.; Farghadani, R.; Namvar, F.; Mohamad, J.; Abdul Kadir, H. Cytotoxic Effects and Anti-Angiogenesis Potential of Pistachio (Pistacia Vera L.) Hulls against MCF-7 Human Breast Cancer Cells. Molecules 2018, 23, 110. [Google Scholar] [CrossRef] [PubMed]
- Reboredo-Rodríguez, P.; González-Barreiro, C.; Cancho-Grande, B.; Simal-Gándara, J.; Giampieri, F.; Forbes-Hernández, T.Y.; Gasparrini, M.; Afrin, S.; Cianciosi, D.; Manna, P.P.; et al. Effect of Pistachio Kernel Extracts in MCF-7 Breast Cancer Cells: Inhibition of Cell Proliferation, Induction of ROS Production, Modulation of Glycolysis and of Mitochondrial Respiration. Journal of Functional Foods 2018, 45, 155–164. [Google Scholar] [CrossRef]
- Bongiorno, D.; Musso, N.; Bonacci, P.G.; Bivona, D.A.; Massimino, M.; Stracquadanio, S.; Bonaccorso, C.; Fortuna, C.G.; Stefani, S. Heteroaryl-Ethylenes as New Lead Compounds in the Fight against High Priority Bacterial Strains. Antibiotics (Basel) 2021, 10, 1034. [Google Scholar] [CrossRef]
- Bonomo, C.; Bonacci, P.G.; Bivona, D.A.; Mirabile, A.; Bongiorno, D.; Nicitra, E.; Marino, A.; Bonaccorso, C.; Consiglio, G.; Fortuna, C.G.; et al. Evaluation of the Effects of Heteroaryl Ethylene Molecules in Combination with Antibiotics: A Preliminary Study on Control Strains. Antibiotics (Basel) 2023, 12, 1308. [Google Scholar] [CrossRef]
- Bivona, D.A.; Mirabile, A.; Bonomo, C.; Bonacci, P.G.; Stracquadanio, S.; Marino, A.; Campanile, F.; Bonaccorso, C.; Fortuna, C.G.; Stefani, S.; et al. Heteroaryl-Ethylenes as New Effective Agents for High Priority Gram-Positive and Gram-Negative Bacterial Clinical Isolates. Antibiotics 2022, 11, 767. [Google Scholar] [CrossRef]
- Consoli, G.M.L.; Maugeri, L.; Musso, N.; Gulino, A.; D’Urso, L.; Bonacci, P.; Buscarino, G.; Forte, G.; Petralia, S. One-Pot Synthesis of Luminescent and Photothermal Carbon Boron-Nitride Quantum Dots Exhibiting Cell Damage Protective Effects. Adv Healthc Mater 2024, 13, e2303692. [Google Scholar] [CrossRef]
- Bonacci, P.G.; Caruso, G.; Scandura, G.; Pandino, C.; Romano, A.; Russo, G.I.; Pethig, R.; Camarda, M.; Musso, N. Impact of Buffer Composition on Biochemical, Morphological and Mechanical Parameters: A Tare before Dielectrophoretic Cell Separation and Isolation. Transl Oncol 2023, 28, 101599. [Google Scholar] [CrossRef]
- Musso, N.; Bonacci, P.G.; Letizia Consoli, G.M.; Maugeri, L.; Terrana, M.; Lanzanò, L.; Longo, E.; Buscarino, G.; Consoli, A.; Petralia, S. Biofriendly Glucose-Derived Carbon Nanodots: GLUT2-Mediated Cell Internalization for an Efficient Targeted Drug Delivery and Light-Triggered Cancer Cell Damage. Journal of Colloid and Interface Science 2025, 696, 137873. [Google Scholar] [CrossRef]
- Fetting, C. The European Green Deal; Vienna, 2020. [Google Scholar]
- Chemicals Strategy - European Commission. Available online: https://environment.ec.europa.eu/strategy/chemicals-strategy_en (accessed on 15 June 2025).
- Chemat, F.; Abert-Vian, M.; Fabiano-Tixier, A.S.; Strube, J.; Uhlenbrock, L.; Gunjevic, V.; Cravotto, G. Green Extraction of Natural Products. Origins, Current Status, and Future Challenges. TrAC Trends in Analytical Chemistry 2019, 118, 248–263. [Google Scholar] [CrossRef]
- Gil-Martín, E.; Forbes-Hernández, T.; Romero, A.; Cianciosi, D.; Giampieri, F.; Battino, M. Influence of the Extraction Method on the Recovery of Bioactive Phenolic Compounds from Food Industry By-Products. Food Chem 2022, 378, 131918. [Google Scholar] [CrossRef]
- Shen, L.; Pang, S.; Zhong, M.; Sun, Y.; Qayum, A.; Liu, Y.; Rashid, A.; Xu, B.; Liang, Q.; Ma, H.; et al. A Comprehensive Review of Ultrasonic Assisted Extraction (UAE) for Bioactive Components: Principles, Advantages, Equipment, and Combined Technologies. Ultrason Sonochem 2023, 101, 106646. [Google Scholar] [CrossRef] [PubMed]
- Schneider, Y.K.; Jørgensen, S.M.; Andersen, J.H.; Hansen, E.H. Qualitative and Quantitative Comparison of Liquid–Liquid Phase Extraction Using Ethyl Acetate and Liquid–Solid Phase Extraction Using Poly-Benzyl-Resin for Natural Products. Applied Sciences 2021, 11, 10241. [Google Scholar] [CrossRef]
- Mendoza-Pérez, R.J.; Álvarez-Olmedo, E.; Vicente, A.; Ronda, F.; Caballero, P.A. Oil Extraction Systems Influence the Techno-Functional and Nutritional Properties of Pistachio Processing By-Products. Foods 2025, 14, 2722. [Google Scholar] [CrossRef]
- Halwatura, L.M.; Aga, D.S. Broad-Range Extraction of Highly Polar to Non-Polar Organic Contaminants for Inclusive Target Analysis and Suspect Screening of Environmental Samples. Science of The Total Environment 2023, 893, 164707. [Google Scholar] [CrossRef]
- Shen, L.; Pang, S.; Zhong, M.; Sun, Y.; Qayum, A.; Liu, Y.; Rashid, A.; Xu, B.; Liang, Q.; Ma, H.; et al. A Comprehensive Review of Ultrasonic Assisted Extraction (UAE) for Bioactive Components: Principles, Advantages, Equipment, and Combined Technologies. Ultrasonics Sonochemistry 2023, 101, 106646. [Google Scholar] [CrossRef] [PubMed]
- Campbell, N.K.; Fitzgerald, H.K.; Fletcher, J.M.; Dunne, A. Plant-Derived Polyphenols Modulate Human Dendritic Cell Metabolism and Immune Function via AMPK-Dependent Induction of Heme Oxygenase-1. Front Immunol 2019, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Wei, P.; Li, Z.; Wei, R.; Li, H.; Li, S. The Role of P62/Nrf2/Keap1 Signaling Pathway in Lead-Induced Neurological Dysfunction. CNS Neurosci Ther 2025, 31, e70566. [Google Scholar] [CrossRef]
- Erlank, H.; Elmann, A.; Kohen, R.; Kanner, J. Polyphenols Activate Nrf2 in Astrocytes via H2O2, Semiquinones, and Quinones. Free Radical Biology and Medicine 2011, 51, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Kanner, J. Polyphenols by Generating H2O2, Affect Cell Redox Signaling, Inhibit PTPs and Activate Nrf2 Axis for Adaptation and Cell Surviving: In Vitro, In Vivo and Human Health. Antioxidants (Basel) 2020, 9, 797. [Google Scholar] [CrossRef]
- Pittala, V.; Vanella, L.; Salerno, L.; Romeo, G.; Marrazzo, A.; Di Giacomo, C.; Sorrenti, V. Effects of Polyphenolic Derivatives on Heme Oxygenase-System in Metabolic Dysfunctions. CMC 2018, 25, 1577–1595. [Google Scholar] [CrossRef]
- Akkus, E.; Yekedüz, E.; Ürün, Y. Sodium-Glucose Cotransporter 2 (SGLT2) as a Potential Biomarker and Target in Papillary Renal Cell Carcinoma. Clin Genitourin Cancer 2025, 23, 102314. [Google Scholar] [CrossRef]
- Musso, N.; Bonacci, P.G.; Letizia Consoli, G.M.; Maugeri, L.; Terrana, M.; Lanzanò, L.; Longo, E.; Buscarino, G.; Consoli, A.; Petralia, S. Biofriendly Glucose-Derived Carbon Nanodots: GLUT2-Mediated Cell Internalization for an Efficient Targeted Drug Delivery and Light-Triggered Cancer Cell Damage. J Colloid Interface Sci 2025, 696, 137873. [Google Scholar] [CrossRef]
- Ahmed, N.; Berridge, M.V. Regulation of Glucose Transport by Interleukin-3 in Growth Factor-Dependent and Oncogene-Transformed Bone Marrow-Derived Cell Lines. Leuk Res 1997, 21, 609–618. [Google Scholar] [CrossRef]
- Landaverde-Mejia, K.; Dufoo-Hurtado, E.; Camacho-Vega, D.; Maldonado-Celis, M.E.; Mendoza-Diaz, S.; Campos-Vega, R. Pistachio (Pistacia Vera L.) Consumption Improves Cognitive Performance and Mood in Overweight Young Adults: A Pilot Study. Food Chem 2024, 457, 140211. [Google Scholar] [CrossRef]
- Rock, C.L.; Zunshine, E.; Nguyen, H.T.; Perez, A.O.; Zoumas, C.; Pakiz, B.; White, M.M. Effects of Pistachio Consumption in a Behavioral Weight Loss Intervention on Weight Change, Cardiometabolic Factors, and Dietary Intake. Nutrients 2020, 12, 2155. [Google Scholar] [CrossRef]
- Xia, K.; Yang, T.; An, L.-Y.; Lin, Y.-Y.; Qi, Y.-X.; Chen, X.-Z.; Sun, D.-L. The Relationship between Pistachio (Pistacia Vera L) Intake and Adiposity: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Medicine (Baltimore) 2020, 99, e21136. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. The Metabolism of Carcinoma Cells1. The Journal of Cancer Research 1925, 9, 148–163. [Google Scholar] [CrossRef]
- Zardo, N.; Will, C.; Santos, A.V.; Galvan, D.; Carasek, E. Exploring HDES as a Sustainable Green Extractant Phase in HF-MMLLE Technique for Assessment of PAHs in Hot Beverages and Evaluation of Potential Dietary Exposure Risks for the Brazilian Population. Food Chem 2025, 493, 145680. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Li, J.; Chen, Y.; Jia, R.; Zeng, X.; Li, Y.; Chen, D.; Li, D. In-Syringe Polypropylene Fiber-Supported Liquid Microextraction Using Natural Deep Eutectic Solvents for the Convenient Analysis of Amphetamine-Type Stimulants in Biological and Environmental Samples. J Chromatogr B Analyt Technol Biomed Life Sci 2025, 1263, 124701. [Google Scholar] [CrossRef] [PubMed]
- Pittala, V.; Vanella, L.; Salerno, L.; Romeo, G.; Marrazzo, A.; Di Giacomo, C.; Sorrenti, V. Effects of Polyphenolic Derivatives on Heme Oxygenase-System in Metabolic Dysfunctions. Curr Med Chem 2018, 25, 1577–1595. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of the green extract and the classic extract extracts on HCT-116 cell viability after 24, 48, and 72 hours of treatment. Cells were treated with the green extract (indigo bars) or the classic extract (blue bars) at a final concentration of 250 ng/μL. Untreated cells (purple bars) were used as controls. Cell viability was assessed via MTT assay and results are expressed as percentage relative to control (set to 100%). Data are presented as mean ± SD (n = 4). Statistical significance was determined by Ordinary one-way ANOVA multiple comparisons test: * p < 0.05; ** p < 0.01; ***p < 0.001; ****p < 0.0001.
Figure 1.
Effect of the green extract and the classic extract extracts on HCT-116 cell viability after 24, 48, and 72 hours of treatment. Cells were treated with the green extract (indigo bars) or the classic extract (blue bars) at a final concentration of 250 ng/μL. Untreated cells (purple bars) were used as controls. Cell viability was assessed via MTT assay and results are expressed as percentage relative to control (set to 100%). Data are presented as mean ± SD (n = 4). Statistical significance was determined by Ordinary one-way ANOVA multiple comparisons test: * p < 0.05; ** p < 0.01; ***p < 0.001; ****p < 0.0001.
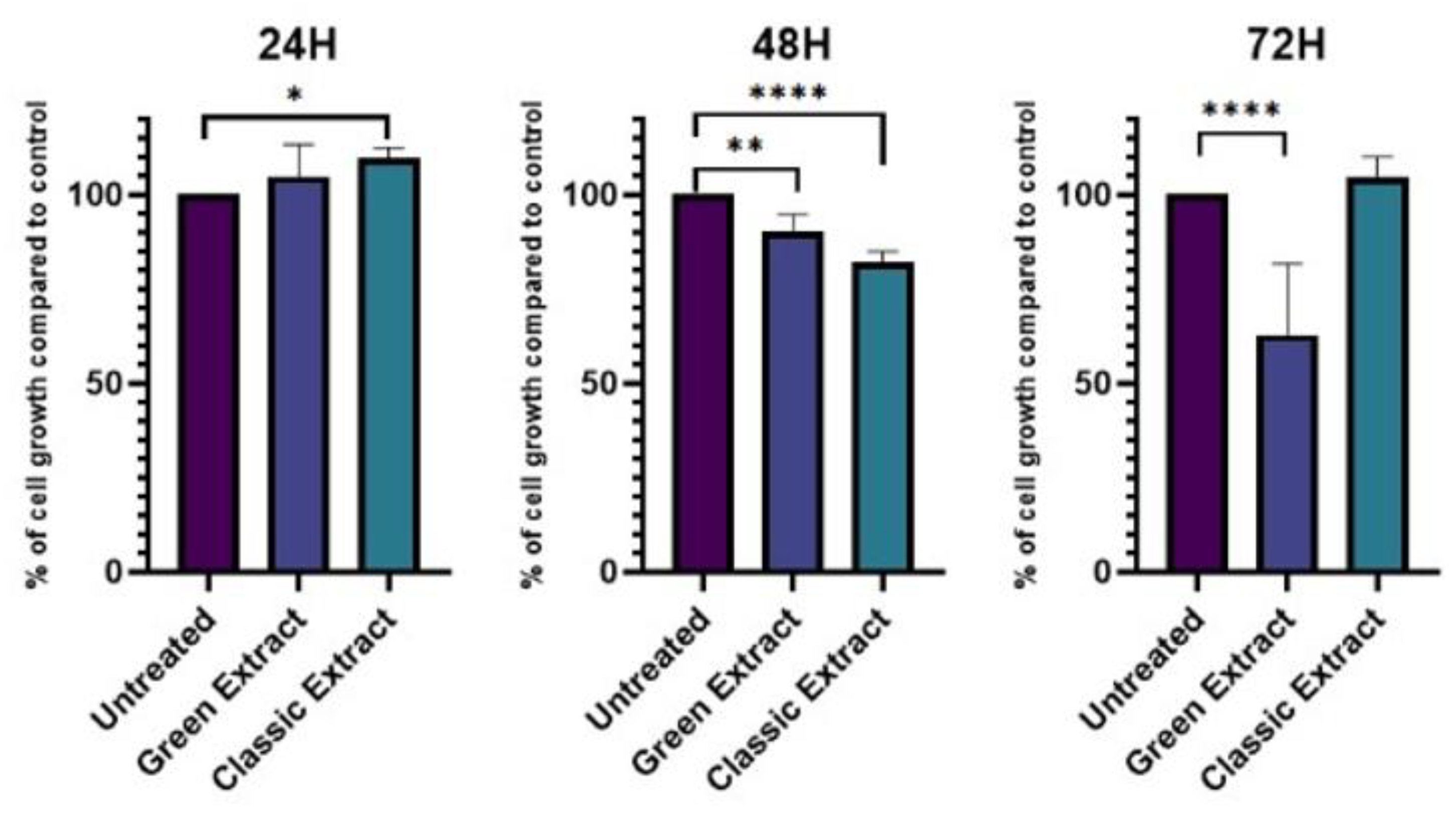
Figure 2.
Effects of pistachio-derived extracts on the expression of IL-6 (Interleukine 6), IL-10, (Interleukine 10) and HMOX-1 (Heme – Oxygenase 1) genes in HCT-116 cells after 24 hours of treatment. Gene expression levels were assessed by RT-qPCR and normalized to the housekeeping gene RPLP0. Results are expressed as fold change relative to untreated control cells (CTRL). Cells were treated with three types of extracts: The one extracted using a green extraction method, and the one extracted using conventional extraction. Statistical significance was determined by Two-stage linear step-up procedure of Benjamini, Krieger and Yekutieli: * p < 0.05; ** p < 0.01; ***p < 0.001; ****p < 0.0001.
Figure 2.
Effects of pistachio-derived extracts on the expression of IL-6 (Interleukine 6), IL-10, (Interleukine 10) and HMOX-1 (Heme – Oxygenase 1) genes in HCT-116 cells after 24 hours of treatment. Gene expression levels were assessed by RT-qPCR and normalized to the housekeeping gene RPLP0. Results are expressed as fold change relative to untreated control cells (CTRL). Cells were treated with three types of extracts: The one extracted using a green extraction method, and the one extracted using conventional extraction. Statistical significance was determined by Two-stage linear step-up procedure of Benjamini, Krieger and Yekutieli: * p < 0.05; ** p < 0.01; ***p < 0.001; ****p < 0.0001.
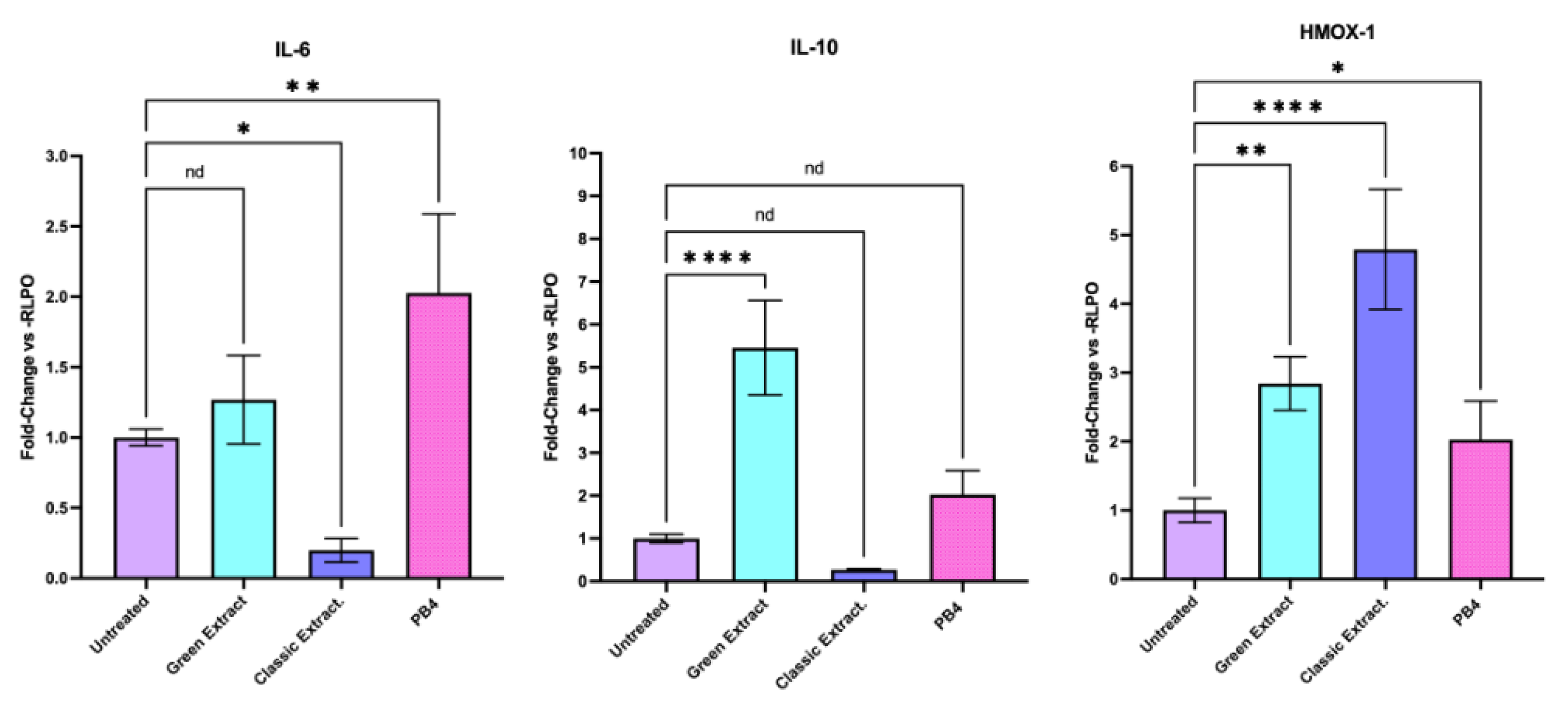
Figure 3.
(Left) Effects of pistachio-derived extracts on the expression of GLUT2 (Solute Carrier Family 2) gene in HCT-116 cells after 24 hours of treatment. Gene expression levels were assessed by RT-qPCR and normalized to the housekeeping gene RPLP0. Results are expressed as fold change relative to untreated control cells (CTRL). Cells were treated with two types of extracts: the one extracted using a green extraction method, and the one extracted using conventional extraction. (Right) The expression of GLUT2 in HCT-116 cells exposed to three different glucose concentrations: Basal (3 g/L), Deprivation (0g/L) and Plus (10 g/L)[21]. Statistical significance was determined by Two-stage linear step-up procedure of Benjamini, Krieger and Yekutieli: * p < 0.05; ** p < 0.01; ***p < 0.001; ****p < 0.0001.
Figure 3.
(Left) Effects of pistachio-derived extracts on the expression of GLUT2 (Solute Carrier Family 2) gene in HCT-116 cells after 24 hours of treatment. Gene expression levels were assessed by RT-qPCR and normalized to the housekeeping gene RPLP0. Results are expressed as fold change relative to untreated control cells (CTRL). Cells were treated with two types of extracts: the one extracted using a green extraction method, and the one extracted using conventional extraction. (Right) The expression of GLUT2 in HCT-116 cells exposed to three different glucose concentrations: Basal (3 g/L), Deprivation (0g/L) and Plus (10 g/L)[21]. Statistical significance was determined by Two-stage linear step-up procedure of Benjamini, Krieger and Yekutieli: * p < 0.05; ** p < 0.01; ***p < 0.001; ****p < 0.0001.
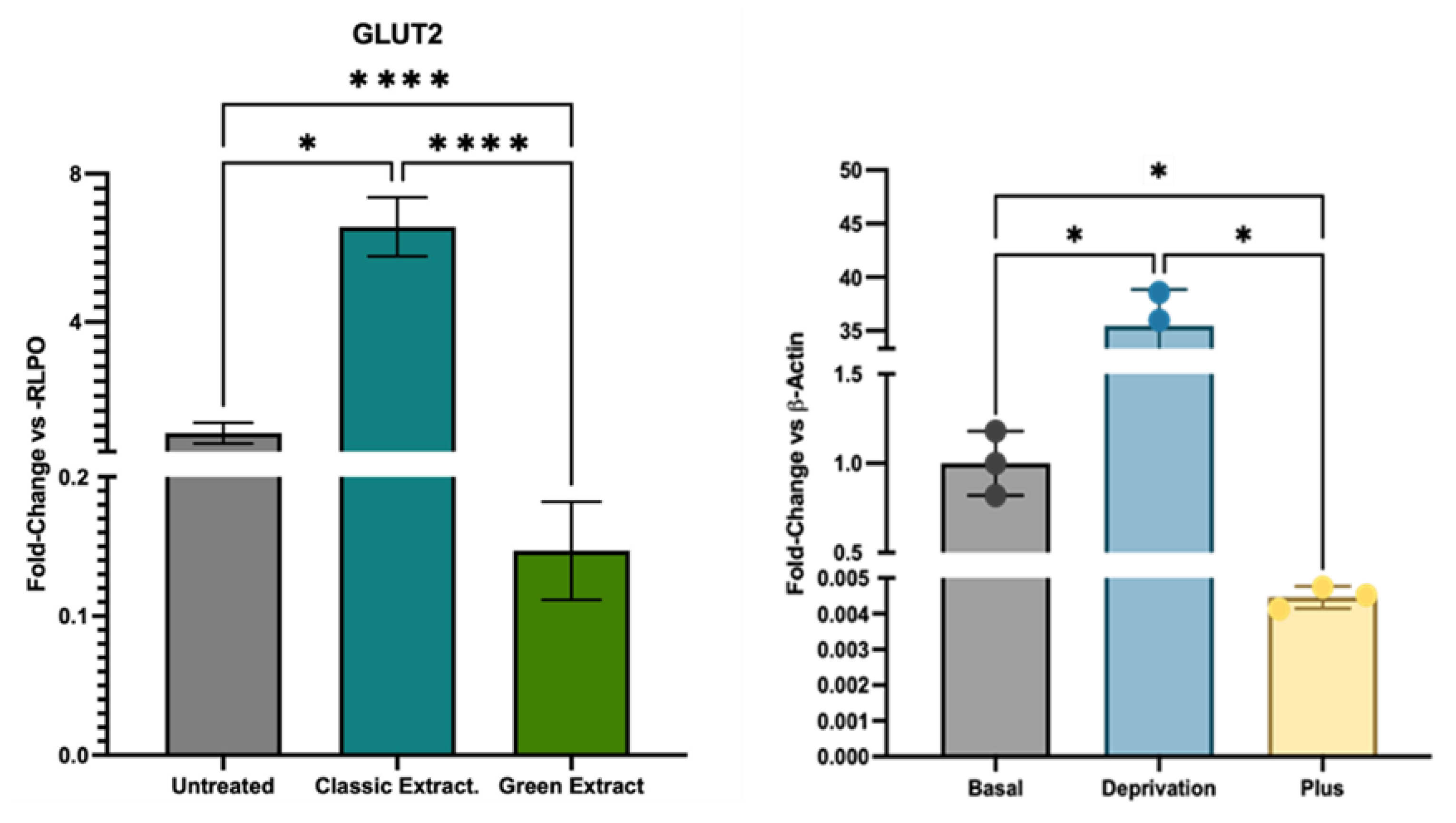

Table 1.
Primers used for RT-qPCR in this study. All primers were ordered from GeneGlobe, QIAGEN (cat. no. 249990).
Table 1.
Primers used for RT-qPCR in this study. All primers were ordered from GeneGlobe, QIAGEN (cat. no. 249990).
| Official Name | Official Symbol | Alternative Titles/Symbols | Detected Transcript | Amplicon Lenght |
|---|---|---|---|---|
| Ribosomal Protein Lateral Stalk Subunit P0 |
RPLP0 | PRLP0; P0; L10E; RPP0; LP0 | NM_053275 NM_001002 |
68 bp |
| Heme Oxygenase 1 | HMOX-1 | HO-1 | NM_002133 | 161 bp |
| Solute Carrier Family 2 Member 2 |
GLUT2 |
GLUT2; Solute Carrier Family 2 Member 2; Glucose Transporter Type 2, Liver |
NM_000340; NM_001278658; NM_001278659 |
120 bp |
| Interleukin 6 | IL-6 | IL-6 | NM_031168 | 128 bp |
| Interleukin 10 | IL-10 | IL-10 | NM_000572 | 112 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



