
Submitted:
30 September 2025
Posted:
01 October 2025
You are already at the latest version
Abstract
Black soldier fly (Hermetia illucens) larvae are a promising source of insect oil, characterized by rapid fatty acid accumulation and a high lauric acid content. This study investigated the effects of dietary black soldier fly oil (BSFO) on muscle quality in largemouth bass (Micropterus salmoides). Experimental diets were formulated to be isonitrogenous, isolipidic, and isophosphoric, with 1.0% and 2.0% BSFO partially replacing soybean oil. A control group received 2.3% soybean oil without BSFO or glycerol monolaurate (GML), while positive controls were supplemented with 0.35% and 0.7% GML. Fish (initial weight: 25.08 ± 0.12 g) were cultured in pond cages for 56 days.Muscle quality and nutritional traits were evaluated, including proximate composition, fatty acid profiles, texture properties, fiber diameter, hydroxyproline content, antioxidant capacity, and expression of genes related to muscle development, atrophy, apoptosis, and mTOR signaling. Compared with the control, the 2.0% BSFO group showed a significant increase in muscle hydroxyproline content (p < 0.05), while GML supplementation led to a significant decrease (p < 0.05). In the 1.0% BSFO group, muscle SFA and MUFA contents were unchanged (p > 0.05), but n-3/n-6 PUFA ratio and HUFA levels were significantly elevated (p < 0.05). The dietary supplementation of BSFO enhanced the levels of high-quality fatty acids in the muscle tissue. Antioxidant capacity was also significantly enhanced in the 1.0% BSFO group (p < 0.05), but reduced in the GML groups (p < 0.05). Texture analysis showed that BSFO significantly improved muscle hardness, elasticity, chewiness, and gumminess (p < 0.05). Gene expression analysis revealed no significant effects of BSFO on genes related to myogenesis(myod、myog)and muscle atrophy(mstn, murf1), or apoptosis-related genes (caspase8, caspase9, caspase3) (p > 0.05); mTOR signaling pathway-related genes (s6k1, akt1) were significantly upregulated in the 2.0% BSFO group (p < 0.05). In contrast, 0.7% GML significantly upregulated genes related to myogenesis(myod, myf5, myog), muscle atrophy(mstn, fbxo32, murf1), and apoptosis (caspase8, caspase9, caspase3) (p < 0.05). In summary, dietary supplementation with 1.0% BSFO effectively enhances muscle quality in largemouth bass without negatively impacting muscle development.
Keywords:
black soldier fly
; insect lipid
; largemouth bass
; flesh quality
; texture property
1. Introduction
Lipids are indispensable in fish nutrition, serving as a vital source of energy and essential nutrients that are fundamental to the growth and health of aquatic species. Traditionally, soybean oil has been a preferred lipid source in commercial fish feed due to its high quality and nutritional value [1]. However, the extensive use of plant-based fats, such as soybean oil, has raised concerns over its competition with human land resources and the associated environmental implications, including deforestation and the excessive application of fertilizers and pesticides. For achieving the sustainable development of aquaculture, it has become important to find new sources of high-quality fats with low-cost and less impact on environment.
Insects have garnered extensive attention from researchers as a potential feed source for some aquatic animals in recent years [2]. Insects have their advantages such as high fertility, fast growth rate, renewable resources and the ability to utilize organic waste. The black soldier fly (Hermitia illucens) is one of the resource insects designated by the Food and Agriculture Organization (FAO) of the United Nations due to its nutritional comprehensiveness, well tolerance and ease of factory farming [3]. Black soldier fly larvae are rich in medium-chain fatty acids, with lauric acid constituting for 21.4-49.3% of total fatty acids [4]. Initially considered a by-product, black soldier fly oil (BSFO) has now attracted attention as a suitable lipid source due to the high lipid content and unique fatty acid profile [5].
Currently, black soldier fly oil has been reported as a fat source for aquatic animal feed. Fawole et al. [6] found that feeding BSFO did not adversely affect the normal performance of juvenile rainbow trout (Oncorhynchus mykiss) and fatty acid deposition in muscle. At the same time, adding BSFO to feed could promot growth, effectively prevent the development of enteritis, and improve the immunity of rainbow trout [5]. Sudha et al. [7] found that BSFO significantly enhanced the expression levels of myod and myog in the white muscle of juvenile striped catfish (Pangasianodon hypophthalmus), thereby promoting muscle development. Furthermore, adding BSFO to feed affects the fatty acid composition of fish because of the high content of lauric acid in BSFO [8,9,10]. So far, BSFO has been less studied in largemouth bass (Micropterus salmoides). Xia et al. [11] found that replacing fish oil with BSFO in largemouth bass feed had no negative effects on growth and decreased liver glycogen content. Conversely, adding BSFO to feed was beneficial to the immunity promotion of largemouth bass.
Largemouth bass (M. salmoides), known for its excellent growth performance, strong resilience, and low disease incidence, has become a key economic freshwater fish species [12]. FAO statistics reveal that global annual production of largemouth bass increased from 621 kilotonnes to 804 kilotonnes between 2020 and 2022 [13]. With the expansion of largemouth bass farming, there is a growing imperative to develop new feed lipid sources. Moreover, as living standards improve, consumer demand for high-quality aquatic products is increasing, making the flesh quality of fish a critical consideration when seeking lipid substitutes.
In this study, we investigated the effects of partially replacing soybean oil with BSFO in the feed on the body composition and texture of largemouth bass. Our findings aim to provide data supporting the substitution of soybean oil with BSFO in feed and contribute to the development of new, low-carbon feed options.
2. Materials and Methods
2.1. Experimental Feed and Groups
Five isonitrogenous and isolipidic feeds with 53% crude protein and 8.6% crude lipid were formulated by using fishmeal and soybean meal as the main protein sources, soybean oil , BSFO and glyceryl monolaurate as the main lipid sources. BSFO free diet was as Control. Experimental diets were formulated to be isonitrogenous, isolipidic, and isophosphoric, with 1.0% and 2.0% BSFO partially replacing soybean oil. The control diet contained 2.3% soybean oil with neither BSFO nor glycerol monolaurate (GML), whereas the positive control diets were supplemented with 0.35% and 0.7% GML. Feed ingredients were thoroughly pulverized to pass through a 60-mesh sieve, full mixing, mixed oil and the right amount of water added to make pellets. The pellets feed were dried at 40 ℃ and stored at -20 ℃ until use. The experimental diet formulation and proximate composition were shown in Table 1.
Black soldier fly larvae were rinsed three times with tap water and ultrapure water respectively, and then dried using vacuum freeze-drying. The dried BSFL were subjected to mechanical pressing to extract oil. The extracted oil was centrifuged at 13,000 rpm for 10 minutes at 40 °C, and the clear supernatant was collected as the BSFO used in this study.
2.2. Experimental Fish and the Feeding Trial
The experimental fish were feed in net cages (1.8 m × 1.5 m × 1.8 m) in a pond covering an area of 10,000 m2. The experimental fish were feed with the diet of the control group for two weeks for acclimation. 600 healthy largemouth bass were randomly divided into 15 net cages (1.8m × 1.5m × 1.8m) with 40 fish per cage, with an initial average body weight of 25.08 ± 0.12 g. During the eight-week culture trial, fish were fed twice daily (06:30 and 17:00) with 3% of body weight. During farming, the water temperature is maintained at 24-35 ℃, dissolved oxygen > 6 mg/L, pH 7.6±0.5, NH4+-N < 0.01 mg/L.
2.3. Sample Collection
After 8-week feeding trial, the fish were fasted for 24 h and anesthetized with an appropriate amount of MS-222. Three fish were taken from each net cage and stored at -20 °C for body component analysis. Nine fish in each net cage were randomly selected for dissection, and muscles above the left lateral line were collected, frozen in liquid nitrogen and stored at -80 °C for biochemical analysis and gene expression analysis. Another 3 fish were dissected, the muscle above the left lateral line were stored in a 4% paraformaldehyde solution at 4 ℃ for histologic section preparation. Three fish were taken from each net cage to remove scales and skin, and the muscle below the dorsal fin and above the left lateral line was cut into 1 cm × 1 cm × 1 cm square muscle blocks for texture analysis.
2.4. Proximate Analysis of Body and Muscle Composition
The moisture, crude protein, crude fat and crude ash was calculated according to National Standards, GB5009.3-2016, GB6432-2018, GB/T14772-2008 and GB5009.4-2016 respectively. The fatty acid composition was determined with an fatty acid profile analyzer (6980N, Agilent Technologies, USA)., and glycogen and hydroxyproline content of the muscle were determined using the A043-1-1 and A030-2-1 commercial assay kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.5. Muscle Antioxidant Capacity
Total superoxide dismutase (T-SOD) and total antioxidant capacity (T-AOC) ain muscle were measured by colorimetric method using the A001-1-2 and A015-1-2 commercial assay kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.6. Texture Properties of Muscle
Six textural parameters including hardness, elasticity, chewiness, cohesiveness, gumminess and resilience were measured using a texture analyzer (Rapid TA+, Shanghai Tengba Instrument Technology, China). The analyzer was operated with the following parameters: P/36R probe diameter of 3.6 mm, test speeds of 5 mm/s, downward pressure distance of 30% of the sample thickness at 4 sec intervalsand trigger force setting of 5 g.
2.7. Histology of Muscle
Muscle samples were dehydrated in ethanol, infiltrated in xylene, embedded in paraffin, cut serially (5 μm thick) and then stained with hematoxylin and eosin. Section were observed using a light microscope (DM500, Leica, German) and photographed. The diameters of the muscle fibers were expressed as the longest axis length using the imaging software (ImageJ, USA).
2.8. RT-qPCR Analysis
Total RNA was extracted from muscle using Trizol reagent. RNA concentration was determined using a micro UV spectrophotometer (NanoDrop 2000, Thrrmo, USA). The total RNA was reverse-transcribed into cDNA with the PrimerScriptTM RT reagent kit with gDNA Eraser (TaKaRa, Dalian, China) and then stored at -20℃. The relative expression levels of gapdh used as an internal reference, and the relative expression levels of the target genes were calculated using the 2-∆∆CTmethod [14]. Primers designed by Primer Premier 5.0 and the sequences were shown in Table 2.
2.9. RT-qPCR Analysis
SPSS 26 was used to statistical analysis. One-way ANOVA was carried out with Duncan’s multiple range test at a significance level of 0.05. All the result are presented as the mean ± s tandard errors (SEM). Bar graphs were produced using Origin 2021 software.
3. Results
3.1. The Effect of Black Soldier Fly Oil Supplementation on the Body and Muscle Composition
The composition of fish body and muscle fed with BSFO replacing different levels of soybean oil was shown in Table 3. Fish fed with different levels of BSFO had no significant impact on crude lipid, crude protein and ash of whole body and muscle of M. salmoides (p > 0.05).
3.2. The Effect of Black Soldier Fly Oil and Glyceryl Monolaurate Supplementation on the Glycogen and Hydroxyproline Contents of Muscle
The effect of black soldier fly oil and glyceryl monolaurate replacing soybean oil on the glycogen and hydroxyproline content of M. salmoides muscle was shown in Table 4. There were no significant effect on glycogen contents in the groups of BSFO and GML compared with the control group (p > 0.05). Hydroxyproline content was significantly increased in the group BSFO2, whereas it was decreased in both GML0.35 and GML0.7 groups (p < 0.05).
3.3. The Effect of Black Soldier Fly Oil Supplementation on the Fatty Acid Composition of Body and Muscle
Table 5 showed the effects on the fatty acid composition of body and muscle of M. salmoides by black soldier fly oil supplementation. C12:0 (lauric acid), C14:0 and ∑SFA in the body were significantly more content in the BSFO supplementation group compared with the control group (p < 0.05), and C20:5n-3 (EPA), C22:6n-3 (DHA), ∑n-3 PUFA and ∑HUFA were significantly lower content in the BSFO groups compared with the control group (p < 0.05 ). The n-3/n-6 PUFA ratio was not significantly different in all groups (p > 0.05).
In the muscle, EPA, HUFA and∑n-3 PUFA were significantly higher content in the BSFO groups compared with the control group (p < 0.05), and the contents of DHA and ∑n-6 PUFA were significantly less than the control group (p < 0.05). The n-3/n-6 PUFA ratio was significantly higher in all BSFO groups than the control group (p < 0.05).
3.4. The Effect of Black Soldier Fly Oil and Glyceryl Monolaurate Supplementation on the Antioxidant Capacity of Muscle
There was no significant difference in T-SOD activity among the groups (p > 0.05) shown in Table 6. The T-AOC in the muscle was not affected by BSFO supplementation (p > 0.05), and was significantly lower in the GML0.35 and GML0.7 groups compared with the control group (p < 0.05). MDA content in the muscle was not affected by BSFO1 supplementation (p > 0.05), and increased significantly in BSFO2, GML0.35 and GML0.7 groups (p < 0.05).
3.5. The Effect of Black Soldier Fly Oil Supplementation on the Texture of Muscle
As shown in Table 7, the hardness, elasticity, chewiness and gumminess of muscle significantly increased compared with the control (p < 0.05), and cohesiveness and resilience were not affected by BSFO supplementation (p > 0.05).
3.6. The Effect of Black Soldier Fly Oil Supplementation on the Histology of Muscle
As shown in Figure 1, compared with the control, the mean diameter of myofibers and the distribution of them in the muscle were not affected by BSFO supplementation. (p > 0.05 ). The histological morphological structure of M. salmoides myofiber was similar in both the soybean oil feed group and the BSFO groups.
3.7. The Effect on the Gene Relative Expression Levels in the Muscle by Fat Source Supplementation
The relative expression levels of myogenesis genes, TOR path-related genes and muscle apoptosis-related genes of M. salmoides were shown in Figure 2. Compared with the control group, the relative expression levels of myf5 related to myog was significantly lower in the BSFO1 group (p < 0.05), and the relative expression levels of fbxo32 related to muscle atrophy was significantly higher in the BSFO2 group (p < 0.05). Adding BSFO to diet reduced the relative expression levels of 4ebp1 significantly (p < 0.05). The relative expression levels of akt1 and s6k1 were significantly higher in the BSFO2 group (p < 0.05). The results showed that the addition of BSFO in the diet had no significant effect on the relative expression levels of apoptosis-related genes (p > 0.05).
Compare with the control group, the relative expression levels of all genes related to myogenesis, TOR path and muscle apoptosis were significantly higher in the GML0.7 group (p < 0.05), and the relative expression levels of fbxo32, s6k1, caspase8 and caspase9 were significantly lower in the GML0.35 group (p < 0.05).
4. Discussion
4.1. Effect on Flesh Quality of Largemouth Bass by BSFO Replacing Soybean Oil
The flesh textural characteristics, pivotal in quality assessment, are typically evaluated using texture profile analysis method [15]. This method quantifies key textural parameters such as hardness, elasticity, chewiness, cohesiveness, gumminess, and resilience, mimicking the human chewing process. These parameters are influenced by several factors, including intramuscular and intermuscular fat content, collagen content in connective tissues, and myofiber diameter and density [16]. Notably, hardness is a critical parameter for consumers, often reflecting the meat perceived value [17].Previous studies on fish have shown that the addition of black soldier fly oil alters the texture of flesh in terms of changes in hardness, elasticity, chewiness and gumminess [18]. In this study, we found that hardness, elasticity, chewiness and gumminess reached the higher values in the BSFO1 group, while they decreased in the BSFO2 group which had a higher degree of substitution compared with the BSFO1 group, but were still higher than the control group. This result is similar to previous discovery in M. salmoides where the addition of BSFO to the diet significantly increased flesh hardness [18]. Among the texture characteristics of flesh, especially hardness, are closely related to muscle lipid content [19]. In the present study, muscle lipid content of M.salmoides did not change significantly when the fish fed with BSFO instead of soybean oil. Previous studies in O. mykiss, Cyprinus carpio var. Jian and M.salmoides have also found that the usage of BSFO in diets could not vary lipid content of muscle [6,8,11].
In addition, the characteristics of fish myofibers influence the quality of the flesh, particularly the hardness, which is directly related to myofiber diameter and density [20]. Previous studies have shown that hardness and chewiness of flesh decreased as myofiber diameter increases and myofiber density decreases [21,22]. However, in this study, the increase in muscle hardness was not attributed to alterations in myofiber diameter, as substituting soybean oil with BSFO did not yield significant changes in either the diameter or density of largemouth bass myofiber. It was contrary to the reported by Xia et al., who found that when BSFO replaced more than 50% of fish oil, there was an increase in the diameter of M.salmoides myofiber; conversely, no significant change occurred when the replacement ratio remained below 50% [11]. It is possible that the difference in results is due to the difference between the fish oil and soybean oil used in the control group. Studies have shown that caspase mediated apoptosis was negatively correlated with myofiber diameter [23]. In this study, fish fed BSFO had no significant impact on the relative expression levels of apoptosis initiator caspase8 and caspase9 and apoptosis effector caspase3. However, the relative expression levels of caspase8, caspase9 and caspase3 was significantly upregulated in the GML0.35 and GML0.7 group. This suggested that lauric acid can induce apoptosis. Interestingly, despite the high lauric acid content in BSFO, its inclusion in fish diets did not exhibit these adverse effects, highlighting the potential of BSFO as a valuable feed component, even with its high concentration of lauric acid.
Furthermore, collagen content also affects the hardness of fish muscles. As a key component of the extracellular matrix, collagen is crucial for sustaining muscle growth and texture in fish [24]. Hydroxyproline, constituting 13.4% of the total amino and imine content in collagen and present in trace amounts in elastin, is absent in other proteins, making it a reliable indicator of collagen levels in muscle tissues [25]. In this study, we found that hydroxyproline content increased in the muscle of M.salmoides fed BSFO instead of soybean oil , which indicated that the increase in collagen amount is a reason of the increase in the flesh hardness.
In order to explore the mechanisms behind the observed changes in the muscle texture of M.salmoides, we examined the expression levels of genes associated with muscle growth. Østbye et al. [26] found a positive correlation between muscle hardness and myogenesis in Salmo salar. The process of fish muscle growth is regulated by a variety of factors, including myogenic regulatory factors (MRFs) and atrophy factors. MRFs are a family of basic helix-loop-helix transcription factors including myod, myf5, mrf4 and myog, and can convert a large number of different cell type into muscle [27]. When the inhibition of myogenesis and protein degradation by atrophy factors exceed protein synthesis, muscle protein loss occurs, leading to a corresponding decrease in muscle hardness [28]. The genes mstn, fbxo32 and MuRF1 are three reliable atrophy sign to indicate muscle protein losing [29,30,31]. After an 8-week feeding trial with dietary BSFO, we assessed its impact on the relative expression levels of myod, myf5, myog, mstn, fbxo32, and MuRF1 in the muscle. Previous studies have shown that some of the trophic factors affected the expression levels of one or more genes in MRFs, such as methionine levels of the dietary for rainbow trout juveniles [32], lysine and histidine levels of the dietary for Oreochromis niloticus [33], plant protein levels of the dietary for Solea senegalensis [34]. Sudha et al. [7] found that replacing fish oil with BSFO increasing the expression levels of genes related to muscle myogenesis in Pangasianodon hypophthalmus while reducing myostatin expression levels. However, another study on replacing fish oil in M.salmoides diets with BSFO found that there were no significant differences in mRNA expression levels of myod1, murf1, myos, myog, myf5 and paxbp-1 [11]. In this study, we replaced soybean oil in largemouth bass diets with BSFO, and found that the relative expression levels of genes related to muscle myogenesis and atrophy were not affected in the group BSFO2 compared with the control group. In the BSFO1 group, the expression level of myf5 decreased significantly, while the expression level of fbxo32 increased significantly, which revealing the primary cause why muscle hardness in BSFO1 group was higher than that in the control group and the BSFO2 group.
In this study, the relative expression levels of genes related to muscle myogenesis and atrophy were up-regulated in the GML0.7 group compare with the control. Despite the substantial lauric acid content in BSFO, the relative expression levels of these genes in the BSFO group were significantly lower than those in the GML group. This suggests that certain components within BSFO may mitigate the adverse effects typically associated with lauric acid, highlighting the potential for BSFO to modulate muscle gene expression in a beneficial manner.
The target of rapamycin (TOR) pathway integrates signals from extracellular and intracellular agents, which can regulate protein synthesis and promote cell survival, proliferation and growth [35]. In the TOR pathway, kinase akt activates TOR target proteins, phosphorylates kinase S6K1 and eukaryotic translation initiation factor 4E-BP1 regulate protein synthesis to promote muscle growth [36]. In the present study, the relative expression levels of atk1 and s6k1 was significantly up-regulated, suggesting that BSFO promotes cell growth and may lead to increased hardness in muscle.
4.2. Effect of Diet BSFO on Fatty Acid Profile of Largemouth Bass
The fatty acid profile in fish is directly affected by the fatty acid composition of the feed [37]. The fatty acid composition of BSFO is predominantly SFA, the majority of which is lauric acid [38]. The results showed that as the level of BSFO supplementation in the feed increased, the content of lauric acid increased in whole body and muscle of largemouth bass. Similar results have been reported in gilthead seabream [9]. In addition, feeding BSFO instead of soybean oil can resulte in an increase of C22:6n-3 (DHA) in muscle in Jian carp [8]. However, the fatty acid profile in rainbow trout was not consistent with them, where the usage of BSFO replacing soybean oil had no significant change in the DHA content of the muscle [6]. In the present study, replacing soybean oil with BSFO did not cause DHA content change in largemouth bass muscle, but significantly elevated C20:5n-3 (EPA) content in muscle, which may be attributed to the fact that lauric acid can be preferentially oxidized, allowing EPA to be better retained in muscle. The result that whole body contains more lauric acid than muscle seems to support this idea. In addition, the n-3/n-6 ratio is an important parameter for assessing the nutrition of flesh. Studies have shown that a high n-3/n-6 ratio is favorable for human health because n-3 PUFA can reduces inflammatory responses, whereas n-6 PUFA causes inflammation contrarily [39]. In the present study, n-3/n-6 levels were also significantly higher in the BSFO1 and BSFO2 groups than in the control group. This is in agreement with the results of previous studies [6,10]. Thus, the addition of BSFO to the diet increased EPA levels and n-3/n-6 ratio in M.salmoides muscle, thereby improved fatty acid quality accordingly.
4.3. Effect of Diet BSFO on Antioxidant Capacity of Largemouth Bass
T-AOC is the total level of various antioxidant macromolecules, antioxidant small molecules and enzymes within a system [40,41]. T-SOD is an antioxidant enzyme that protects cells from peroxidative damage [42]. In the present study, replacing soybean oil with BSFO had no significant effect on both T-AOC and T-SOD activities in muscle. The results showed that inclusion of BSFO up to 2% in the diet had no significant effect on the antioxidant capacity of largemouth bass. Similar results have been reported, such as in the juvenile yellow catfish, BSFO as a substitute for soybean oil has no significant effect on the antioxidant capacity [43]. However, another study found that the addition of BSFO to largemouth bass diets resulted in an increase in T-SOD activities in the liver, suggesting that BSFO improves the antioxidant capacity of largemouth bass [11]. Because different studies have produced conflicting results, to investigate how BSFO affects the antioxidant capacity of largemouth bass, we fed largemouth bass using GML partially substitution of soybean oil. The results showed that the total antioxidant capacity of largemouth bass was significantly reduced. However, this may not be a universal phenomenon. For example, the study in hybrid grouper indicated GML increased the activity of SOD [44]. In Trachinotus ovatus, 0.15% GML significantly increased total antioxidant capacity and superoxide dismutase activities [45]. Similarly, the dietary addition of GML significantly increased the SOD activity in the juvenile grouper [46]. The different results may be due to the different effects of GML on different species of fish. In the present study, total antioxidant capacity was significantly lower in the GML groups than in the control, whereas the BSFO groups remained comparable to the control, suggesting that certain components in BSFO might mitigate the GML induced impairment of antioxidant capacity in largemouth bass.
5. Conclusions
This paper objective estimated the effects of dietary BSFO supplementation on the whole fish body composition, muscle composition, and fatty acid content, texture characteristics, diameter of muscle fibers, collagen content, antioxidant capacity of muscle, and mRNA expression levels of related genes involved in muscle generation, atrophy and apoptosis of largemouth bass. Our results showed that the addition of BSFO to the diet increased muscle hardness, enhanced flesh quality, also increased EPA levels and n-3/n-6 ratio in largemouth bass muscle and improved fatty acid quality. BSFO had no effect on the antioxidant capacity. In summary, it is feasible to use BSFO in the feed for largemouth bass.
Author Contributions
Conceptualization, P.W.; methodology, P.W.; software, Z.W. (Zichuan Wang); formal analysis, Z.W. (Zichuan Wang), Y.C., W.Y., Z.W. (Zeting Wang) and Y.K.; investigation, W.Y., Z.W. (Zeting Wang) and Y.K.; resources, P.W.; data curation, Z.W. (Zichuan Wang), Y.C., W.Y. and P.W.; writing—original draft preparation, Z.W. (Zichuan Wang), Y.C.; writing—review and editing, Z.W. (Zichuan Wang), Y.C. and P.W.; supervision, P.W., C.C. and Y.Y.; project administration, P.W., C.C. and Y.Y.; funding acquisition, P.W. and Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Natural Science Foundation of the Higher Education Institutions of Jiangsu Province (20KJA240001); the Foundation of ReProtein Biotechnology (Suzhou) Co., Ltd. ( H211372), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Institutional Review Board Statement
This study was conducted according to the SOPs of the Provincial Aquatic Animal Nutrition Key Laboratory of Soochow University and approved by the Animal Welfare Ethics Committee of Soochow University (Approval No.SUDA20250320A12).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors thank all the students and staff who contributed to and supported the entire study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Peng, S.M.; Chen, L.Q.; Qin, J.G.; Hou, J.L.; Yu, N.; Long, Z.Q.; Ye, J.Y.; Sun, X.J. Effects of replacement of dietary fish oil by soybean oil on growth performance and liver biochemical composition in juvenile black seabream, Acanthopagrus schlegeli. Aquaculture 2008, 276, 154–161. [Google Scholar] [CrossRef]
- Moon, S.J.; Lee, J.W. Current views on insect feed and its future. Entomological Research 2015, 45, 283–285. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: establishing optimal feeding rates. Waste Management & Research 2009, 27, 603–610. [Google Scholar] [CrossRef]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens) —Possibilities and limitations for modification through diet. Waste Management 2020, 102, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Fawole, F.J.; Romano, N.; Hossain, M.S.; Labh, S.N.; Overturf, K.; Small, B.C. Insect (black soldier fly, Hermetia illucens) meal supplementation prevents the soybean meal-induced intestinal enteritis in rainbow trout and health benefits of using insect oil. Fish & Shellfish Immunology 2021, 109, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Fawole, F.J.; Labh, S.N.; Hossain, M.S.; Overturf, K.; Small, B.C.; Welker, T.L.; Hardy, R.W.; Kumar, V. Insect (black soldier fly larvae) oil as a potential substitute for fish or soy oil in the fish meal-based diet of juvenile rainbow trout (Oncorhynchus mykiss). Animal Nutrition 2021, 7, 1360–1370. [Google Scholar] [CrossRef] [PubMed]
- Sudha, C.; Ahilan, B.; Felix, N.; Uma, A.; Chidambaram, P.; Prabu, E. Replacement of fish oil with black soldier fly larvae oil and vegetable oils: Effects of growth, whole-body fatty acid profile, digestive enzyme activity, haemato-biochemical responses and muscle growth-related gene expression of juvenile striped catfish, Pangasianodon hypophthalmus. Aquaculture Research 2022, 53, 3097–3111. [Google Scholar] [CrossRef]
- Li, S.L.; Ji, H.; Zhang, B.X.; Tian, J.J.; Zhou, J.S.; Yu, H.B. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance, body composition, tissue fatty acid composition and lipid deposition in juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Moutinho, S.; Oliva-Teles, A.; Pulido-Rodríguez, L.; Parisi, G.; Magalhaes, R.; Monroig, O.; Peres, H. Effects of black soldier fly (Hermetia illucens) larvae oil on fillet quality and nutritional traits of gilthead seabream. Aquaculture 2024, 579. [Google Scholar] [CrossRef]
- Goda, A.M.A.; El-Haroun, E.; Nazmi, H.; Van Doan, H.; Aboseif, A.M.; Taha, M.K.S.; Abou Shabana, N.M. Black soldier fly oil-based diets enriched in lauric acid enhance growth, hematological indices, and fatty acid profiles of Nile tilapia, Oreochromis niloticus fry. Aquaculture Reports 2024, 37. [Google Scholar] [CrossRef]
- Xia, M.; Hu, Z.C.; Wang, G.; Yuan, X.T.; Ji, H.; Sun, J.; Yu, H.B. Evaluation of black soldier fly (Hermetia illucens) larvae oil as a potential lipid source for largemouth bass (Micropterus salmoides) : Growth performance, myofiber development and glucolipid metabolism. Animal Feed Science and Technology 2024, 318. [Google Scholar] [CrossRef]
- Wang, D.; Yao, H.; Li, Y.H.; Xu, Y.J.; Ma, X.F.; Wang, H.P. Global diversity and genetic landscape of natural populations and hatchery stocks of largemouth bass Micropterus salmoides across American and Asian regions. Scientific Reports 2019, 9. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2024, /: Available online: https, 1428.
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research 2001, 29. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.H.; Zhang, Z.; Weng, M.; Shen, Y.A.; Lai, W.C.; Hao, T.T.; Yao, C.W.; Bu, X.Y.; Du, J.L.; Li, Y.R.; et al. Glycerol monolaurate improved intestinal barrier, antioxidant capacity, inflammatory response and microbiota dysbiosis in large yellow croaker (Larimichthys crocea) fed with high soybean oil diets. Fish & Shellfish Immunology 2023, 141. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.T. Technology integration and innovation to improve the edible quality of freshwater cultured fish. Feed Industry 2022, 43, 1–11. [Google Scholar] [CrossRef]
- de Huidobro, F.R.; Miguel, E.; Blázquez, B.; Onega, E. A comparison between two methods (Warner-Bratzler and texture profile analysis) for testing either raw meat or cooked meat. Meat Science 2005, 69, 527–536. [Google Scholar] [CrossRef]
- Yuan, H.L.; Hu, J.R.; Li, X.C.; Sun, Q.X.; Tan, X.H.; You, C.H.; Dong, Y.W.; Huang, Y.H.; Zhou, M. Dietary black soldier fly oil enhances growth performance, flesh quality, and health status of largemouth bass (Micropterus salmoides). Animal Nutrition 2024, 18. [Google Scholar] [CrossRef] [PubMed]
- Boleman, S.J.; Boleman, S.L.; Miller, R.K.; Taylor, J.F.; Cross, H.R.; Wheeler, T.L.; Koohmaraie, M.; Shackelford, S.D.; Miller, M.F.; West, R.L.; et al. Consumer evaluation of beef of known categories of tenderness. Journal of Animal Science 1997, 75, 1521–1524. [Google Scholar] [CrossRef]
- Sampath, W.; Zhang, Y.; Liu, J.H.; Yang, M.X.; Zhang, W.B.; Mai, K.S. Dietary taurine improves muscle growth and texture characteristics in juvenile turbot (Scophthalmus maximus). Aquaculture Reports 2020, 17. [Google Scholar] [CrossRef]
- Hu, Z.C.; Li, H.D.; Liu, S.; Xue, R.R.; Sun, J.; Ji, H. Assessment of black soldier fly (Hermetia illucens) larvae meal as a potential substitute for soybean meal on growth performance and flesh quality of grass carp Ctenopharyngodon idellus. Animal Nutrition 2023, 14, 425–449. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Cornet, J.; Donnay-Moreno, C.; Gouygou, J.P.; Bergé, J.P.; Bacelar, M.; Escórcio, C.; Rocha, E.; Malhao, F.; Cardinal, M. Quality differences of gilthead sea bream from distinct production systems in Southern Europe: Intensive, integrated, semi-intensive or extensive systems. Food Control 2011, 22, 708–717. [Google Scholar] [CrossRef]
- Stratos, I.; Li, Z.D.; Rotter, R.; Herlyn, P.; Mittlmeier, T.; Vollmar, B. Inhibition of caspase mediated apoptosis restores muscle function after crush injury in rat skeletal muscle. Apoptosis 2012, 17, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Lin, J.L.; Liu, X.; Yuan, Y.Y.; Liu, G.Q.; Ye, X.K. Effects of temperature on muscle growth and collagen deposition in zebrafish (Danio rerio). Aquaculture Reports 2022, 22. [Google Scholar] [CrossRef]
- Burjanadze, T.V. Hydroxyproline Content and Location in Relation to Collagen Thermal-Stability. Biopolymers 1979, 18, 931–938. [Google Scholar] [CrossRef]
- Ostbye, T.K.K.; Ruyter, B.; Standal, I.B.; Stien, L.H.; Bahuaud, D.; Dessen, J.E.; Latif, M.S.; Fyhn-Terjesen, B.; Rorvik, K.A.; Morkore, T. Functional amino acids stimulate muscle development and improve fillet texture of Atlantic salmon. Aquaculture Nutrition 2018, 24, 14–26. [Google Scholar] [CrossRef]
- Weintraub, H. The Myod Family and Myogenesis-Redundancy, Networks, and Thresholds. Cell 1993, 75, 1241–1244. [Google Scholar] [CrossRef]
- Kitajima, Y.; Yoshioka, K.; Suzuki, N. The ubiquitin–proteasome system in regulation of the skeletal muscle homeostasis and atrophy: from basic science to disorders. The Journal of Physiological Sciences 2020, 70, 40. [Google Scholar] [CrossRef]
- Rodriguez, J.; Vernus, B.; Chelh, I.; Cassar-Malek, I.; Gabillard, J.C.; Sassi, A.H.; Seiliez, I.; Picard, B.; Bonnieu, A. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cellular and Molecular Life Sciences 2014, 71, 4361–4371. [Google Scholar] [CrossRef]
- Coelho, P.D.; Guarnier, F.A.; Figueiredo, L.B.; Zaramela, L.S.; Pacini, E.S.A.; Godinho, R.O.; Gomes, M.D. Identification of potential target genes associated with the reversion of androgen-dependent skeletal muscle atrophy. Archives of Biochemistry and Biophysics 2019, 663, 173–182. [Google Scholar] [CrossRef]
- Kakareka, K.; Waddell, D. Characterizing the Role of Muscle-Specific RING Finger 1 (MuRF1) as a Regulator of Atrophy-Induced Gene Expression. Faseb Journal 2017, 31, 31. [Google Scholar] [CrossRef]
- Alami-Durante, H.; Cluzeaud, M.; Bazin, D.; Schrama, J.W.; Saravanan, S.; Geurden, I. Muscle growth mechanisms in response to isoenergetic changes in dietary non-protein energy source at low and high protein levels in juvenile rainbow trout. Comparative Biochemistry and Physiology a-Molecular & Integrative Physiology 2019, 230, 91–99. [Google Scholar] [CrossRef]
- Richter, B.L.; Silva, T.S.D.; Michelato, M.; Marinho, M.T.; Gonçalves, G.S.; Furuya, W.M. Combination of lysine and histidine improves growth performance, expression of muscle growth-related genes and fillet quality of grow-out Nile tilapia. Aquaculture Nutrition 2021, 27, 568–580. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Cabral, E.M.; Sousa, V.; Cunha, L.; Fernandes, J.M.O. Plant protein blends in diets for Senegalese sole affect skeletal muscle growth, flesh texture and the expression of related genes. Aquaculture 2016, 453, 77–85. [Google Scholar] [CrossRef]
- Thomas, G.; Hall, M.N. TOR signalling and control of cell growth. Current Opinion in Cell Biology 1997, 9, 782–787. [Google Scholar] [CrossRef]
- Lushchak, O.; Strilbytska, O.; Piskovatska, V.; Storey, K.B.; Koliada, A.; Vaiserman, A. The role of the TOR pathway in mediating the link between nutrition and longevity. Mechanisms of Ageing and Development 2017, 164, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, I.; Biasibetti, E.; et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. Journal of Animal Science and Biotechnology 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Animal Feed Science and Technology 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Fabrikov, D.; Barroso, F.G.; Sánchez-Muros, M.J.; Hidalgo, M.C.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F.; Guil-Guerrero, J.L. Effect of feeding with insect meal diet on the fatty acid compositions of sea bream (Sparus aurata), tench (Tinca tinca) and rainbow trout (Oncorhynchus mykiss) fillets. Aquaculture 2021, 545. [Google Scholar] [CrossRef]
- Gyan, W.R.; Yang, Q.H.; Tan, B.P.; Dong, X.H.; Chi, S.Y.; Liu, H.Y.; Zhang, S. Effects of replacing fishmeal with dietary dried distillers grains with solubles on growth, serum biochemical indices, antioxidative functions, and disease resistance for Litopenaeus vannamei juveniles. Aquaculture Reports 2021, 21. [Google Scholar] [CrossRef]
- Lin, H.X.; Yang, Q.H.; Wang, A.J.; Wang, J.X.; Tan, B.P.; Ray, G.W.; Dong, X.H.; Chi, S.Y.; Liu, H.Y.; Zhang, S. Effects of fish meal under different storage conditions on growth, serum biochemical indices and antioxidant capacity for juvenile grouper Epinephelus coioides. Aquaculture Nutrition 2021, 27, 723–733. [Google Scholar] [CrossRef]
- Anuar, N.S.; Shafie, S.A.; Maznan, M.A.F.; Zin, N.; Azmi, N.A.S.; Raoof, R.A.; Myrzakozha, D.; Samsulrizal, N. Lauric acid improves hormonal profiles, antioxidant properties, sperm quality and histomorphometric changes in testis and epididymis of streptozotocin-induced diabetic infertility rats. Toxicology and Applied Pharmacology 2023, 470. [Google Scholar] [CrossRef]
- Hu, J.R.; Yi, J.C.; Wang, G.X.; Mo, W.Y.; Huang, Y.H. Effects of dietary soybean oil replace with black soldier fly larvae oil on growth performance, plasma biochemical indexes and liver lipid droplets of juvenile yellow catfish. Acta Hydrobiologica Sinica 2020, 44, 717–727. [Google Scholar]
- Li, X.H.; Yi, Y.M.; Wu, J.H.; Yang, Q.H.; Tan, B.P.; Chi, S.Y. Effects of Plant-Derived Glycerol Monolaurate (GML) Additive on the Antioxidant Capacity, Anti-Inflammatory Ability, Muscle Nutritional Value, and Intestinal Flora of Hybrid Grouper (Epinephelus fuscoguttatus♀ x Epinephelus lanceolatus♂). Metabolites 2022, 12. [Google Scholar] [CrossRef]
- Lin, H.X.; Tan, B.P.; Yang, Q.H.; Chi, S.Y.; Wei, H.J.; Wu, Y.C.; Ray, G.W.; Yohana, M.A. Effects of Dietary Glycerol Monolaurate on Growth, Antioxidant Capacity and Lipid Metabolism in Cage-Farmed Pompano (Trachinotus ovatus) Juveniles. Frontiers in Marine Science 2022, 9. [Google Scholar] [CrossRef]
- Li, X.H.; Zhu, D.W.J.; Mao, M.L.; Wu, J.W.; Yang, Q.H.; Tan, B.P.; Chi, S.Y. Glycerol Monolaurate Alleviates Oxidative Stress and Intestinal Flora Imbalance Caused by Salinity Changes for Juvenile Grouper. Metabolites 2022, 12. [Google Scholar] [CrossRef]
Figure 1.
Figure 1. The effect of black soldier fly oil replacing soybean oil on the histology of muscle. (A), (B), (C) Transversal section of the white muscle of fish fed different experimental diets. Hematoxylin-eosin staining. Photomicrographs (magnification 100×) and scale bar (100μm); (D) Average diameter of myofibers; (E) the distribution of myofiber diameters.
Figure 1.
Figure 1. The effect of black soldier fly oil replacing soybean oil on the histology of muscle. (A), (B), (C) Transversal section of the white muscle of fish fed different experimental diets. Hematoxylin-eosin staining. Photomicrographs (magnification 100×) and scale bar (100μm); (D) Average diameter of myofibers; (E) the distribution of myofiber diameters.
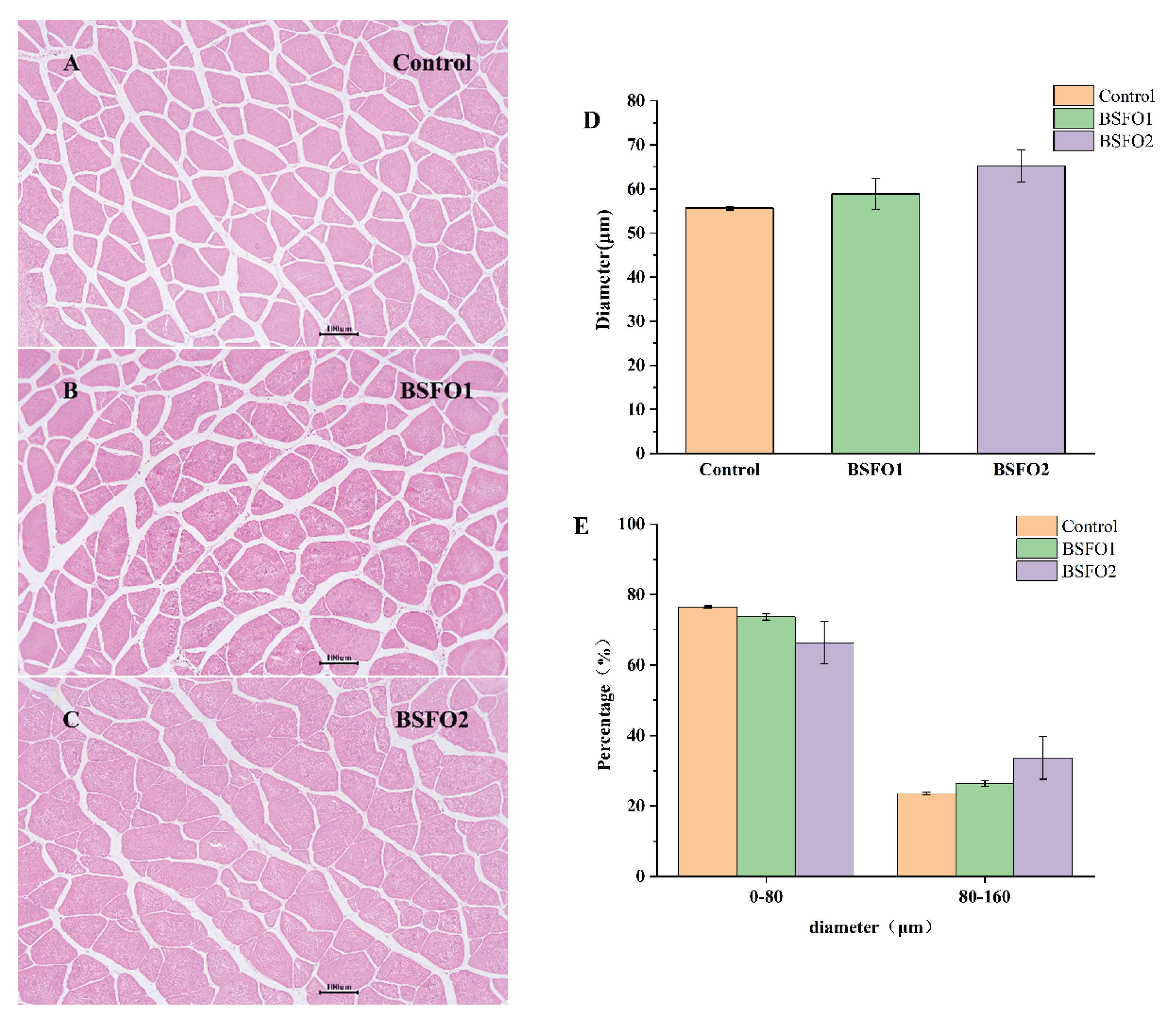
Figure 2.
Effects of black soldier oil replacing soybean oil on the regulation of myogenesis, TOR path-related genes and apoptosis-related genes of largemouth bass. (A) myogenesis regulation-related genes (B) TOR path-related genes (C) apoptosis-related genes.
Figure 2.
Effects of black soldier oil replacing soybean oil on the regulation of myogenesis, TOR path-related genes and apoptosis-related genes of largemouth bass. (A) myogenesis regulation-related genes (B) TOR path-related genes (C) apoptosis-related genes.
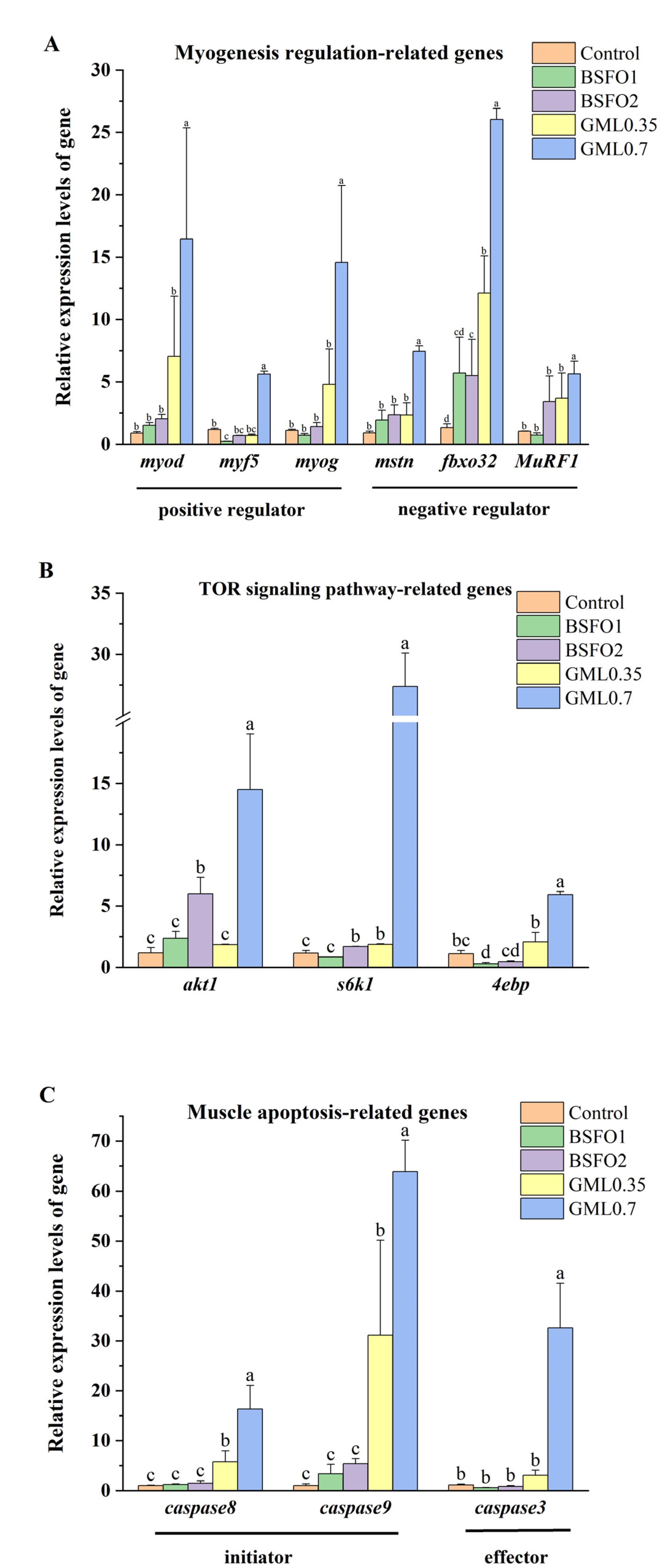

Table 1.
Largemouth bass experimental diets composition (g/kg) and nutrient composition.
| Items | Control | BSFO1 | BSFO2 | GML0.35 | GML0.7 |
|---|---|---|---|---|---|
| Tapioca | 100 | 100 | 100 | 100 | 100 |
| Full-Fat Rice Bran | 116.5 | 116.5 | 116.5 | 116.5 | 116.5 |
| Cotton meal | 80 | 80 | 80 | 80 | 80 |
| Corn gluten meal | 35 | 35 | 35 | 35 | 35 |
| Fish meal | 500 | 500 | 500 | 500 | 500 |
| Hemoglobin powder | 30 | 30 | 30 | 30 | 30 |
| Chicken meal | 90 | 90 | 90 | 90 | 90 |
| Black soldier fly oil | 0 | 10 | 20 | 0 | 0 |
| Soybean oil | 23 | 13 | 3 | 19.5 | 16 |
| glyceryl monolaurate | 0 | 0 | 0 | 3.5 | 7 |
| Ca (H2PO4) 2 | 15 | 15 | 15 | 15 | 15 |
| Premix1 | 10 | 10 | 10 | 10 | 10 |
| Ruiantai2 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Proximate analysis (%, DM) | |||||
| Moisture | 10.67 | 6.56 | 5.51 | 11.23 | 10.09 |
| Crude protein | 53.30 | 53.40 | 53.12 | 53.37 | 53.22 |
| Crude protein | 8.66 | 8.64 | 8.77 | 8.66 | 8.70 |
| Ash | 13.51 | 13.33 | 13.43 | 13.39 | 13.40 |
1Premix contained the following per kilogram of feed:vitamin A 8mg,vitamin B1 18mg,vitamin B2 8mg,vitamin B6 12mg,vitamin B12 0.02mg,vitamin C 300mg,vitamin D3 3mg,vitamin K3 5mg, folic acid 5 mg, pantothenic acid 25 mg, niacin 25 mg, inositol 100 mg,Mg 96mg,Fe 64mg,Zn 19mg,Mn 13mg,Cu 2.5mg,I 0.021mg,Se 0.07mg,Co 0.016mg,K 0.05mg. 2Ruiantai is natutal plant-based additives.
Table 2.
The sequences of gene primers used for RT-qPCR.
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| myf5 | GAGGAGGACGAGCATGTCAG | GAGGTGCAACGTCTCAAAGC |
| myod | GCGACTAAGCAAGGTGAACG | CTGCAGGGACTCGATGTAGC |
| myog | GAGTTGGGGTGACAGGAACA | TCTGGTTTGGGTTCATCAGG |
| mstn | TGATTGCTTTGGGTCCAGTC | CCTTCATTCGCAGTTTGCTC |
| fbxo32 | GACATGGCTGCCAAGAAGAG | TGAAGGCCTCTCCCAGTGTA |
| MuRF1 | TGGACGACACGTGTAAGCTG | GTGGACACCTTCTGCTGCTC |
| caspase7 | CCATGGTGAAGAGGGAATGA | CGGACCCGAATCTGTCTGTA |
| caspase8 | GAGGGGACAAAGAGGTGGAG | GAGCCTGTGGAAGTGTTTCG |
| caspase9 | TCGGGCCTTTCCATTATTTC | AACAACCTGAGTGGGTGCTG |
| akt1 | AGGACGCTACTACGCCATGA | CCGCTCCTCTGAGAACACAC |
| s6k1 | GGCTCATCCCTTCTTTCGAC | GGCTTGATTCGCACTCTCAC |
| 4ebp1 | GACTGCCAGAAGACCACTGC | CAGCAGGAACTTTCGGTCAT |
| gapdh | AAGGGTGGTGCCAAGAGAGT | AGTCTTCTGAGTGGCGGTGA |
Table 3.
Effects of black soldier fly oil supplementation on body and muscle composition (dry weight, %).
Table 3.
Effects of black soldier fly oil supplementation on body and muscle composition (dry weight, %).
| Items | Control | BSFO1 | BSFO2 |
|---|---|---|---|
| Whole fish body | |||
| Crude lipid (%) | 20.71±0.98 | 23.06±1.67 | 24.59±1.84 |
| Crude protein (%) | 61.12±1.12 | 59.96±2.52 | 58.68±2.24 |
| Ash (%) | 15.29±0.72 | 13.95±0.46 | 13.95±0.12 |
| Muscle | |||
| Crude lipid (%) | 16.51±0.83 | 17.04±1.06 | 19.80±1.75 |
| Crude protein (%) | 80.09±0.91 | 79.49±2.72 | 79.09±1.94 |
| Ash (Ash,%) | 5.87±0.27 | 5.90±0.12 | 6.27±0.12 |
Control, BSFO1 and BSFO2 represent diets with BSFO inclusion levels of 0%, 1%, and 2%, respectively. All data are expressed as mean ± SEM.
Table 4.
Effect of black soldier fly oil and glyceryl monolaurate supplementation on the glycogen and hydroxyproline content of muscle.
Table 4.
Effect of black soldier fly oil and glyceryl monolaurate supplementation on the glycogen and hydroxyproline content of muscle.
| Items | Control | BSFO1 | BSFO2 | GML0.35 | GML0.7 |
|---|---|---|---|---|---|
| Glycogen (mg/g) | 1.06±0.08 | 1.11±0.08 | 1.1±0.16 | 1.09±0.02 | 1.06±0.04 |
| Hydroxyproline (μg/ml) | 0.56±0.01b | 0.6±0.02b | 0.76±0.05a | 0.41±0.02c | 0.41±0.02c |
Control, BSFO1 and BSFO2 represent diets with BSFO inclusion levels of 0%, 1%, and 2%, respectively. GML0.35 and GML0.7 represent diets with GML inclusion levels of 0.35% and 0.7% respectively. All data are expressed as mean ± SEM. Means in the same row with different superscripts are significantly different (p < 0.05).
Table 5.
Effect of black soldier fly oil supplementation on the fatty acid composition of whole fish and muscle (% total fatty acid).
Table 5.
Effect of black soldier fly oil supplementation on the fatty acid composition of whole fish and muscle (% total fatty acid).
| Items | Body | Muscle | ||||
|---|---|---|---|---|---|---|
| Fatty acid | Control | BSFO1 | BSFO2 | Control | BSFO1 | BSFO2 |
| C12:0 | 0.18±0.06c | 4.47±0.1b | 6.23±0.46a | 0.14±0c | 2.93±0.22b | 4.84±0.33a |
| C13:0 | 0.09±0a | 0.09±0a | 0.06±0b | 0.04±0a | 0.03±0b | 0.02±0b |
| C14:0 | 5.43±0.11b | 6.59±0.14a | 6.36±0.19a | 3.24±0.01c | 4.07±0.08b | 4.42±0.18a |
| C15:0 | 0.59±0.02a | 0.53±0.03b | 0.42±0.03c | 0.34±0a | 0.3±0.01b | 0.29±0b |
| C16:0 | 35.51±0.75a | 34±0.5ab | 32.64±0.86b | 25.71±0.04a | 23.72±0.63b | 23.62±0.05b |
| C17:0 | 0.59±0.02 | 0.53±0.05 | 0.49±0.05 | 0.35±0a | 0.29±0.01c | 0.32±0b |
| C18:0 | 7.03±0.02a | 6.38±0.27b | 6.04±0.28b | 5.36±0.02a | 4.12±0.04b | 4.23±0.18b |
| C21:0 | 0.14±0.03a | 0.1±0.02ab | 0.07±0.01b | 0.48±0.05 | 0.41±0.04 | 0.43±0.01 |
| C23:0 | 0.43±0.03a | 0.3±0.04b | 0.24±0.04b | 0.4±0.06a | 0.1±0.01b | 0.11±0.01b |
| ∑SFA | 50.01±0.73b | 53.05±0.22a | 52.61±0.77a | 36.43±0.03b | 36.37±0.43b | 38.64±0.34a |
| C16:1n-7 | 6.9±0.12 | 6.96±0.31 | 6.67±0.26 | 5.37±0c | 5.85±0.24a | 5.45±0.17ab |
| C18:1n-9 | 33.97±0.17ab | 32.63±1b | 34.51±0.72a | 29.03±0.1a | 26.44±0.92b | 27.09±0.2b |
| C20:1n-9 | 0.39±0.02a | 0.31±0.01b | 0.25±0.01c | 0.28±0.04a | 0.19±0.01b | 0.18±0b |
| C24:1n-9 | 0.18±0.03 | 0.2±0.1 | 0.17±0.03 | 5.45±0.03b | 7.67±0.25a | 7.88±0.29a |
| ∑MUFA | 41.5±0.1 | 40.19±1.03 | 41.7±0.46 | 40.3±0.14 | 40.34±0.79 | 40.8±0.31 |
| C18:2n-6 | 6.18±0.72a | 4.95±0.82ab | 4.08±0.39b | 18.38±0.02a | 16.91±0.72b | 14.93±0.02c |
| C18:3n-6 | 0.15±0.02 | 0.11±0.04 | 0.08±0.02 | 1.33±0.03a | 1.45±0.1a | 1.16±0.01b |
| C20:2n-6 | 0.06±0.01a | 0.04±0ab | 0.03±0b | 0.04±0.01a | 0.04±0ab | 0.03±0b |
| C20:3n-6 | 0.04±0 | 0.03±0.01 | 0.02±0 | 0.2±0.01b | 0.35±0.02a | 0.44±0.07a |
| C20:4n-6 | 0.21±0.01a | 0.14±0.02b | 0.09±0.02c | 0.14±0.02 | 0.14±0.01 | 0.14±0.01 |
| ∑n-6 PUFA | 6.64±0.75 | 5.28±0.88 | 4.3±0.4 | 20.1±0.01a | 18.89±0.83b | 16.7±0.04c |
| C18:3n-3 | 1.23±0.06a | 1.13±0.03ab | 1.06±0.09b | 0.97±0.04a | 0.78±0.04b | 0.91±0.03a |
| C20:3n-3 | 0.2±0.05a | 0.13±0.02b | 0.1±0.01b | 1.07±0.01b | 1.42±0.18a | 1.11±0.08b |
| C20:5n-3 (EPA) | 0.16±0.03a | 0.07±0.03b | 0.1±0.02ab | 1.02±0.12c | 2.08±0.09a | 1.72±0.03b |
| C22:6n-3 (DHA) | 0.26±0.06a | 0.14±0b | 0.12±0.02b | 0.11±0.01 | 0.12±0.02 | 0.12±0.01 |
| ∑n-3 PUFA | 1.85±0.09a | 1.48±0.01b | 1.38±0.12b | 3.17±0.09c | 4.4±0.28a | 3.86±0.02b |
| ∑PUFA | 8.48±0.81a | 6.76±0.88ab | 5.69±0.34b | 23.27±0.11a | 23.29±1.11a | 20.56±0.04b |
| ∑HUFA | 0.82±0.1a | 0.51±0.02b | 0.43±0.01b | 2.54±0.05c | 4.11±0.27a | 3.54±0.1b |
| n-3/n-6 PUFA | 0.28±0.02 | 0.29±0.05 | 0.33±0.05 | 0.16±0b | 0.23±0a | 0.23±0a |
All data are expressed as mean ± SEM. Means in the same row with different superscripts are significantly different (p < 0.05).
Table 6.
Effects of black soldier fly oil and glyceryl monolaurate supplementation on the antioxidant capacity of muscle.
Table 6.
Effects of black soldier fly oil and glyceryl monolaurate supplementation on the antioxidant capacity of muscle.
| Items | Control | BSFO1 | BSFO2 | GML0.35 | GML0.7 |
|---|---|---|---|---|---|
| T-SOD (U/mgprot) | 33.97±0.89 | 33.67±3.05 | 31.43±1.94 | 33.31±1.85 | 31.34±1.12 |
| T-AOC (U/mgprot) | 2.63±0.12a | 2.7±0.27a | 2.95±0.24a | 1.76±0.18b | 1.94±0.11b |
All data are expressed as mean ± SEM. Means in the same row with different superscripts are significantly different (p < 0.05).
Table 7.
Texture of largemouth bass muscle by partially substituting soybean oil with black soldier fly oil.
Table 7.
Texture of largemouth bass muscle by partially substituting soybean oil with black soldier fly oil.
| Items | Control | BSFO1 | BSFO2 |
|---|---|---|---|
| Hardness,g | 218.88±24.03b | 448.18±46.62a | 351.87±33.88a |
| Elasticity,mm | 0.58±0.01b | 0.64±0.01a | 0.62±0.01a |
| Chewiness,mJ | 79.55±11.86b | 178.73±18.88a | 142.16±17.82a |
| Gummeness,g | 135.83±18.11b | 276.26±27.27a | 226.89±24.68a |
| Cohesiveness | 0.62±0.02 | 0.67±0.01 | 0.64±0.03 |
| Resilience | 0.47±0.02 | 0.5±0.01 | 0.48±0.02 |
All data are expressed as mean ± SEM. Means in the same row with different superscripts are significantly different (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



