
Submitted:
12 September 2025
Posted:
19 September 2025
You are already at the latest version
Abstract
Chromium-tanned leather waste presents a significant environmental challenge due to its resistance to microbial degradation and potential to release toxic chromium compounds. This study aimed to evaluate the biodegradability of wet blue leather through thermo-philic composting without the addition of exogenous microorganisms, and to characterise the associated microbial communities. Composting trials were conducted in 2 L and 40 L Dewar vessels under controlled thermophilic conditions. Gravimetric analysis, scanning electron microscopy (SEM), high-throughput sequencing, and microbial culturing were applied. The small-scale system achieved a 38.94% mass loss, whereas the large-scale composting resulted in the complete disintegration of leather into fragments smaller than 1 cm after approximately 40 days. Metagenomic analysis revealed distinct microbial communities in leather compost, enriched with genera such as Sinibacillus, Bdellovibrio, and Thermomyces, which are known for their thermotolerant enzymatic activity. SEM images confirmed bacterial colonisation and biofilm formation on the leather surface. Cultivable strains with metal tolerance and proteolytic potential were also isolated. These findings demonstrate that composting under thermophilic conditions can effectively degrade chromium-tanned leather. The study also provides a basis for the development of targeted microbial treatments or biotechnological strategies to reduce the environmental impact of the leather industry and support circular bioeconomy approaches in waste management.
Keywords:
chromium‐tanned leather
; composting
; biodegradation
; thermophilic microorganisms
; microbial communities
; biofilms
1. Introduction
The management of solid leather waste, particularly from chromium-tanned (wet blue) leather, represents a persistent challenge for both the leather industry and environmental authorities [1]. Leather production generates large volumes of organic waste [2], with hides and skins subjected to tanning processes that confer durability and resistance to microbial degradation but simultaneously hinder their reintegration into biological cycles [3]. This waste primarily originates from the tanning stage, which consumes significant amounts of water and energy and involves the use of various chemicals [4]. One of the main environmental concerns is the use of chromium salts in tanning, as trivalent chromium Cr(III) can oxidise into its hexavalent form Cr(VI), which is highly toxic [5]. This transformation poses a serious environmental hazard, particularly when waste is mismanaged or improperly disposed of, leading to potential contamination of soil and water bodies [6].
Chromium(III) salts are widely used in tanning due to their affinity for cross-linking with collagen [7].
Although various techniques for recycling and resource recovery from leather waste are being investigated, including the development of polymer composites [8,9] and the extraction of chromium or collagen for reuse in industrial or biomaterial applications [10], these approaches remain at an early stage of development and have not yet been implemented on an industrial scale [11,12]. Consequently, conventional disposal methods, such as landfilling or incineration, continue to be used, despite raising serious environmental and regulatory concerns.
Composting is a well-established biological treatment method for organic waste, relying on aerobic microbial metabolism and thermophilic conditions to convert substrates into stable humic products [14]. However, its application to chromium-tanned leather waste remains largely unexplored, mainly due to the presumed toxicity of chromium salts and the structural complexity of the tanned collagen matrix [15, 16]. The tanning process involves the cross-linking of collagen fibres with trivalent chromium, which enhances durability but significantly reduces biodegradability by inhibiting enzymatic activity and microbial colonisation [17,18]. While the partial biodegradability of untanned or vegetable-tanned leather has been demonstrated under composting or anaerobic digestion [19,20,21], the microbial processes specifically involved in the degradation of chromium-tanned leather remain poorly understood [22]. Some chemical and physical treatments, including acid, alkaline, and enzymatic hydrolysis, sometimes assisted by ultrasound, have been explored for chromium recovery; however, their efficiency and scalability remain limited [23]. In parallel, the chemical reactivity of chromium-tanned leather has been studied through its dye adsorption capacity, providing insights into the mass transfer mechanisms during degradation [24]. Additionally, archaeological evidence of microbial decay in vegetable-tanned leather provides valid reference points, although the behaviour of chromium-tanned leather under similar conditions remains insufficiently characterised [25].
In line with the growing interest in circular economy strategies for treatment of tanned leather waste [26,27,28], recent efforts have focused on understanding the microbial and biochemical mechanisms that enable its biodegradation under controlled conditions. Microbial consortia in composting play a pivotal role in degrading complex organic materials into simpler compounds, adapting to environmental stressors, and harbouring enzymatic potential for degradation processes [29]. These consortia comprise diverse microorganisms, including bacteria, fungi, and archaea, each contributing to the breakdown of organic matter during the composting phases [30]. Composting involves a mesophilic phase followed by a thermophilic phase, each dominated by distinct microbial communities [31]. Mesophiles initiate the degradation of simple compounds, while thermophiles take over at higher temperatures to break down more complex materials [32]. Microbial consortia exhibit resilience and adaptability to environmental stressors, such as metal pollution, by developing tolerance mechanisms [33]. These mechanisms include the formation of efflux systems and enzymatic detoxification, which enable the consortia to adapt and survive in contaminated environments, demonstrating the consortium’s potential for environmental remediation [34]. Moreover, the role of biofilm formation and extracellular polymeric substances in enhancing microbial adhesion, enzymatic synergy, and metal tolerance has become increasingly recognised [35]. Furthermore, the simplification of microbial consortia through strategies such as enrichment and dilution can maintain or even enhance their degradation capabilities, emphasising the potential for developing engineered consortia for specific applications in waste management and bioremediation [36].
Within this context, the present study aims to address two main objectives. First, it seeks to establish and validate two thermophilic composting systems (large and small scale) to evaluate the degradation of wet blue leather, utilising a combination of gravimetric analysis and scanning electron microscopy (SEM) to assess structural and morphological alterations in the leather matrix. Second, it seeks to characterise the bacterial and fungal taxa associated with the composting process, using high-throughput sequencing and the isolation of cultivable strains to describe the microbial communities involved in leather degradation. The microbial profiles of composted leather were compared to those of control compost to determine whether distinct consortia emerge in response to the presence of chromium-tanned substrates. By integrating structural, molecular, and microbiological approaches, this work contributes to a better understanding of the biological potential for leather waste valorisation under thermophilic composting conditions. It also provides foundational knowledge for the development of targeted microbial treatments or biotechnological strategies aimed at mitigating the environmental impact of the leather industry and advancing sustainable waste management solutions aligned with circular bioeconomy principles.
2. Results
2.1. Temperature Monitoring and Leather Degradation
Temperature Monitoring and Process Control
In the small-scale composting system (2 L Dewar vessels), thermophilic conditions (≥50 °C) were achieved within 48 hours, with multiple peaks exceeding 60 °C (Figure S1). However, temperature fluctuations were frequent, with several drops below the 50 °C threshold, requiring five reinoculations at approximately 160, 288, 576, 720 and 864 hours to sustain microbial activity. In contrast, the large-scale composting system (40 L Dewar vessel) demonstrated enhanced thermal stability, with temperatures exceeding 70 °C and remaining elevated for extended periods (Figure S2). Only two reinoculations were required during this assay, occurring at approximately 384 and 672 hours, when the temperature dropped below 55 °C.
2.2. Microbial Characterisation: Bacterial Community Composition
2.2.1. Rarefaction, Alpha and Beta Diversity
Rarefaction curves reached saturation across all samples, indicating that sequencing depth was sufficient to capture the bacterial diversity present (Figure S3). Alpha diversity analysis at the amplicon sequence variant (ASV) level showed that compost control samples exhibited substantially higher diversity than leather compost samples. Specifically, observed ASV richness exceeded 680 in control samples (ine19 and ine20), while leather compost samples (ine17 and ine18) showed values below 400 (Figures S4 and S5). Similarly, the Shannon and Simpson indices were consistently higher in the control compost, with Shannon values above 5.1 and Simpson values near 0.99, compared to values below 4.5 and 0.97, respectively, in the leather compost. Beta diversity analysis, using Bray–Curtis dissimilarity and Principal Coordinates Analysis (PCoA), revealed clear compositional separation between sample types in both composting systems (Figures S6 and S7). In the small-scale system, control samples (ine19, ine20) clustered tightly on the right side of Axis 1 (97.7% of explained variance), while leather compost samples (ine17, ine18) grouped on the left. A similar pattern was observed in the large-scale system: control samples (ine21–ine23) clustered on the left side of Axis 1 (65% variance), and leather compost samples (ine24–ine26) formed a distinct cluster on the right, with strong intra-group similarity in both cases. Differential abundance analysis between the small-scale samples (ine17–ine20) revealed that compost control samples (ine19 and ine20) contained significantly higher abundance in 124 bacterial genera. In contrast, only 49 genera were more abundant in the leather compost samples (ine17 and ine18). Overall, compost control samples contained 196 more genera than leather compost samples.
2.2.2. Taxonomic Distribution at Phylum and Genus Levels
In samples ine17 to ine20, the taxonomic classification at the phylum level revealed high similarity across all replicates. The most abundant phyla across both control and leather compost samples were Firmicutes, Proteobacteria, and Bacteroidota (Figure 1; Figure S8).
At the genus level, more evident differences between control compost and leather compost samples were observed (Figure 2; Figure S9). Puia was more abundant in leather compost samples, whereas Ruminofilibacter dominated the control compost and was nearly absent in leather compost.
Genera such as Sinibacillus, Proteiniborus, Desulfotomaculum, Chelativorans, and Bdellovibrio were notably associated with leather compost samples, while Ruminofilibacter [37] was more frequent in the control group.
In samples ine21 to ine26, taxonomic profiles at the phylum level also showed substantial similarity between replicates. Firmicutes and Actinobacteria were generally more abundant in leather compost samples, while Bacteroidota appeared more frequently in control compost (Figure 3; Figure S10).
At the genus level, more distinct differences between control and leather compost samples were again evident (Figure 4; Figure S11). Genera such as Novibacillus, Aeribacillus, and Thermobifida, previously described in high-temperature composting contexts [38,39,40], were detected exclusively in leather compost samples. In contrast, genera like Ruminofilibacter, also present in the small-scale control compost samples, were dominant in the control group and absent in the leather samples. Galbibacter, Brumimicrobium, and Providencia were statistically more abundant in the leather compost, whereas Ruminiclostridium, Petrimonas, and Herbinix were statistically more abundant in the control compost.
2.3. Microbiota Characterisation: Fungal Community Dynamics
2.3.1. Rarefaction, Alpha and Beta Diversity
Rarefaction curves at the Operational Taxonomic Unit (OTU) level reached saturation across all samples, indicating that sequencing depth was sufficient to capture fungal diversity (Figure S12). Alpha diversity analysis, based on ASV-level metrics, revealed variable patterns among sample types. In the small-scale system using the 2 L Dewar vessel (samples ine17 to ine20), observed ASV richness was higher in the control compost samples (ine17 and ine18), while Shannon and Simpson indices were higher in the leather compost samples (ine19 and ine20) (Figure S13). In the large-scale 40 L Dewar vessel (samples ine21 to ine26), observed richness was similar across all samples. However, Shannon and Simpson indices remained consistently higher in the leather compost group compared to the control (Figure S14). Beta diversity analysis, using Bray–Curtis dissimilarity and PCoA, revealed a clear separation between leather compost and control compost samples in both composting systems, with strong similarity among replicates within each group (Figures S15 and S16).
2.3.2. Taxonomic Distribution at Phylum and Genus Levels
In samples ine17 to ine20, taxonomic classification at the phylum level revealed Ascomycota as the dominant phylum across all samples (Figure 5; Figure S17). However, control compost samples (ine17, ine18) showed a higher relative abundance of Basidiomycota compared to leather compost samples (ine19, ine20), which exhibited a greater proportion of unidentified phyla.
At the genus level, Mycothermus was the most abundant genus across both control and leather compost samples (Figure 6; Figure S18). Other genera showed differential presence depending on the sample type. Chrysosporium was more frequently detected in control compost samples, while an unclassified genus from the family Chaetomiaceae appeared predominantly in leather compost samples.
Genera more abundant in leather compost samples included Thermomyces and Remersonia. In contrast, Scutellinia and Thelebolus were more frequently detected in control compost samples. In samples ine21 to ine26, Ascomycota was the predominant phylum, representing more than 95% of relative abundance in all cases (Figure 7; Figure S19).
At the genus level, notable differences were observed between sample types. Mycothermus was dominant in control compost, whereas Melanocarpus was predominant in leather compost samples (Figure 8; Figure S20). The control sample corresponds to the initial time point of the composting process, while the leather compost samples represent the final state. Additional genus-level assignments are available in the supplementary material. Melanocarpus and Thermomyces were primarily detected in leather compost samples, while Remersonia and Dipodascus were more frequently found in the control group.
2.4. Cultivable Bacterial and Fungal Species Identification
After the 44-day composting period, samples were taken from 2L and 40L composting vessels and cultured. The identified strains are listed in Table 1.
2.5. Scanning Electron Microscopy (SEM) & Gravimetric Analysis: Leather Analysis
Gravimetric analysis in the 2L system indicated a 38.94% mass loss of the leather after 44 days (Figure 9). In the 40L setup, leather degradation was so advanced that no substantial fragments (>1 cm) were recovered, precluding gravimetric analysis and suggesting nearly complete degradation (~100%). This improvement is attributed to bioaugmentation with mature compost from the 2L system.
SEM observations were performed on residual fragments (>1cm) of degraded leather recovered from the 40L setup composting process. As shown in Figure 10, the characteristic fibrous structure of leather was preserved, confirming that the fragments corresponded to wet blue leather.
More advanced signs of microbial colonisation and surface disruption are evident in Figure 11 and Figure 12. At higher magnifications (×3500 to ×7000), the leather matrix appears heavily colonised by bacterial cells, with widespread presence of biofilms and extracellular polymeric substances (EPS). The EPS is visible as amorphous material bridging fibre gaps and embedding microbial cells. Microbial morphologies consistent with cocci and bacilli are clearly visible, indicating a multispecies colonisation. In some areas, matrix detachment, surface irregularity, and fibre separation are observed, indicating progressive biodegradation.
3. Discussion
One of the key findings is the significant structural degradation observed in leather fragments exposed to the large-scale composting system. While complete disintegration was only achieved in this system, small pieces (<1 cm) still allowed for microbial identification and SEM analysis. The clear colonisation of these fragments by bacteria and the presence of extensive biofilms confirm the active involvement of microbial consortia in leather degradation [41]. The EPS not only indicate biofilm maturity but also suggests a potential mechanism for chromium tolerance, as EPS have been described as efficient agents in heavy metal binding and flocculation [35].
The cultivation and identification of bacterial and fungal strains isolated from the degraded leather revealed a high prevalence of thermotolerant and stress-resistant genera. These include Bacillus [42,43], Paenibacillus, Aneurinibacillus, Ureibacillus, Aeribacillus, and Chelatococcus, many of which are known for their keratinolytic or proteolytic potential and have been previously detected in high-temperature composting environments. The isolation of Thermomyces lanuginosus, a fungus with high enzymatic activity at elevated temperatures, supports its proposed role in degrading recalcitrant substrates, such as collagen and residual tanning agents [44,45].
At the community level, bacterial and fungal sequencing data confirmed that composting of leather induces a significant shift in microbial composition and diversity. The bacterial richness and diversity were markedly reduced in leather compost samples compared to controls, consistent with the selective pressure imposed by chromium and collagen-rich substrates. Nevertheless, several genera such as Sinibacillus, Desulfotomaculum, Chelativorans, and Bdellovibrio were enriched in the leather compost environment, many of which have been reported in composts containing organic nitrogen and/or metal-contaminated matrices [46,47,48,49].
Differences were observed between the genera present in the leather compost samples and those in the control compost. Among the genera detected exclusively in the leather-degraded samples were Novibacillus, Aeribacillus, and Thermobifida, all of which have been previously described as thermophilic organisms involved in degradation processes under high-temperature composting conditions [38,39,40]. The presence of these genera supports the adaptive capacity of microbial communities in thermophilic composting environments, as well as their potential role in breaking down recalcitrant materials, such as collagen. In contrast, the genus Ruminofilibacter was detected only in control compost samples and was absent from those containing leather. This genus has been described as an efficient degrader of lignocellulose and has been isolated from anaerobic digesters fed with manure[37]. Its absence in the leather compost samples may indicate sensitivity to the presence of tanned waste or limited adaptation to the specific substrates present in this type of compost. Notably, Ruminofilibacter was also identified in the 2 L control compost, further reinforcing its specificity to specific composting environments. The fungal community also exhibited notable shifts, with Melanocarpus, Thermomyces, and Remersonia being characteristic of leather compost samples. These genera are frequently described in thermophilic composting and are known producers of thermostable hydrolases [50]. Their presence correlates with the observed morphological damage in SEM imaging, where fibre disruption and the loss of the periodic collagen structure were evident.
Among the cultivable strains identified in this study, several species belong to genera previously associated with biodegradation and bioremediation activities. For example, Rhodococcus rhodochrous has demonstrated a high capacity to adsorb heavy metal ions such as cadmium (II), lead (II), nickel (II), cobalt (II), and hexavalent chromium (VI) through extracellular polymeric substances [51]. Bacillus pumilus has been described as a bacterium capable of tolerating elevated concentrations of copper, zinc, and lead [52]. Similarly, Pseudomonas aeruginosa is a well-studied species known for its ability to remove heavy metals from contaminated environments [53,54]. At the same time, Bacillus subtilis has been reported to contribute to the decontamination of water polluted with heavy metals [55]. Acinetobacter species have demonstrated remarkable capabilities to degrade fluorides and heavy metals, such as nickel and chromium, with several studies confirming their role in chromium bioremediation [56,57,58]. From a biodegradation perspective, thermotolerant fungal and bacterial genera such as Thermomyces have been highlighted for their production of thermostable degrading enzymes [45, 59]. Other genera like Ochrobactrum ciceri have shown high potential for the bioremediation of nickel and lead [60], while Pseudoxanthomonas has been recognised for its ability to degrade cellulose and lignocellulose [61]. SEM observations revealed progressive matrix deterioration in leather samples, consistent with microbial enzymatic action. Images showed characteristic detachment of fibre bundles, amorphous regions lacking structural order, and extensive biofilm coverage. These results corroborate previous findings that the mechanical integrity of collagen can be disrupted by sustained microbial activity, particularly in the presence of extracellular enzymes and acidic metabolic byproducts. Gravimetric analysis, although limited to the small-scale system due to complete degradation in the large-scale setup, provides further support for microbial degradation. The variable mass loss among samples suggests heterogeneous microbial dynamics, potentially influenced by initial colonization, leather accessibility, and local physicochemical gradients within the compost matrix.
4. Materials and Methods
4.1. Composting Leather Biodegradation: Small- and Large-Scale Approaches
Biodegradation assays were performed using wet blue leather, a semi-processed material obtained through chromium tanning, which imparts a characteristic blue colour and enhances mechanical and thermal stability. This type of leather retains high moisture content and requires further processing for end-use applications [22, 62, 63]. Two composting systems were employed: small-scale composting (using 2L Dewar vessels) and large-scale composting (using a 40L Dewar vessel). For the small-scale setup, two parallel vessels were prepared, each containing a mixture of hay (40%), plant residues (30%), and manure (30%) to support microbial activity and provide a balanced carbon-to-nitrogen ratio. One vessel served as a control, while the second included three 7 × 1.5 cm leather samples, shown in Figure 13B. The initial weights of the leather samples were determined using an ultra-precision balance (Table S1). For the large-scale setup, a mixture of hay (40%), manure (20%) and mature compost from the small-scale system (10%) is used to enhance microbial activity. Three larger wet blue leather samples shown in Figure 13A were composted. Initial weights were determined using an ultra-precision balance (Table S1). The composting process lasted 44 days (1,056 hours) and was performed under thermophilic conditions (≥50 °C), with temperature continuously monitored throughout the period. Reinoculation and aeration were carried out only when the temperature dropped below 55 ºC. This involved removing the composted material from the vessel, adding fresh plant residues, hay, and manure, and thoroughly mixing to restore microbial activity and oxygen availability.
4.2. Metagenomic DNA Extraction and Sequencing
After 44 days of composting, a total of 10 samples (ine17 to ine26, see Table 2) were collected for DNA extraction. These included both bulk compost material and material scraped from the surface of leather samples, obtained from the 2 L and 40 L composting systems. For leather-associated samples, the surface of the leather was gently scraped using a sterile scalpel to recover solid material containing microbial biomass. All samples were homogenised and resuspended in the lysis buffer provided in the NZY Soil gDNA Isolation Kit (NZYtech, Lisboa, Portugal). DNA extraction was then performed according to the manufacturer’s protocol, which includes chemical lysis, mechanical disruption using bead-beating, and purification via silica membrane-based spin columns [64]. Metagenomic sequencing was conducted on the Illumina MiSeq platform, focusing on the 16S rRNA gene for bacterial identification [65] and the ITS regions of the 23S rRNA gene for fungal identification [66].
4.3. Bioinformatics and Statistical Analysis
Amplicon libraries targeting the bacterial 16S rRNA and fungal ITS regions were prepared according to Illumina’s standard protocol and sequenced on an Illumina MiSeq platform (2×300 bp). The complete description of the amplification and library preparation protocol can be found in [67]. The raw sequences generated by Illumina were imported into the Qiime2 bioinformatics tool [68] to perform an initial quality control of the sequences with DADA2. Taxonomic assignment of each amplicon sequence variant (ASV), defined at a 99.9% sequence similarity, was performed using the classify-Sklearn module in combination with the SILVA v138 database [69] for bacteria and UNITE v8.2 for fungi. Statistical and microbial ecology analyses were performed using various R software packages, including Phyloseq [70] and Vegan [71]. Alpha diversity was measured using the richness, Shannon diversity index, and Simpson diversity index. To determine significant differences in the relative abundances of taxa, the DESeq2 test [72] was used. Beta diversity was assessed using Principal Coordinates Analysis (PCoA), based on a Bray-Curtis dissimilarity matrix [73]. PCoA is an ordination method used to evaluate the similarity of microbial communities. The PERMANOVA test was used to determine statistically significant differences between groups [74].
4.4. Cultivable Bacterial and Fungal Species Identification
In addition to metagenomic analysis, compost samples were plated on various growth media TSB media (Tryptic Soy Broth) (Condalab, Madrid, Spain) 3% and 1.5% agar, R2A media [peptone 0.1%; yeast extract 0.05%; dextrose 0.05%; starch 0.05%; K2HPO4 0.03%; Mg2SO4·7H2O 0.005%; sodium pyruvate 0.03%], NAI media (Nutrient Agar; peptone 1%; meat extract 0.5%; NaCl 0.5%; 1.5% agar; pH 7.2) and YM media (Yeast Mold; yeast extract 0.3%; malt extract 0.3%; soybean peptone 0.5%; glucose 1%; agar 1%) to identify cultivable bacterial and fungal strains. Identifications were made by partial sequencing of the 16S rRNA gene and using the NCBI online bioinformatics tool Blast [75] to obtain the closest strains.
4.5. Scanning Electron Microscopy (SEM) and Gravimetric Analysis: Leather Analysis
At the end of the composting period (44 days), leather samples from the small-scale system (2 L Dewar vessels) were recovered and subjected to gravimetric analysis. Recovered fragments were photographed and then washed sequentially with 10% sodium dodecyl sulfate (SDS), 70% ethanol, and distilled water to remove adherent microbial biomass. The samples were then air-dried under controlled laboratory conditions until they reached a constant weight. Final dry weights were measured using an ultra-precision balance. Gravimetric degradation was expressed as the percentage of mass loss relative to the initial dry weight of each sample before composting (see Table S1 in the Supplementary Material). In contrast, leather samples from the large-scale system (a 40-L Dewar vessel) were not suitable for gravimetric analysis due to advanced degradation; only fragmented pieces smaller than 1 cm could be recovered. These remains were instead analysed using Scanning Electron Microscopy (SEM), following the methodology described by Vyskočilová et al. [76], to assess surface morphology, structural degradation, and potential biofilm formation.
5. Conclusions
This study demonstrates the biodegradability of wet blue leather under thermophilic composting conditions without the addition of exogenous microorganisms. The composting system, using Dewar vessels, achieved significant degradation performance. In the small-scale system, an average mass loss of 38.94% was recorded, while the large-scale composting at temperatures above 70 °C for approximately 40 days resulted in the complete disintegration of leather into fragments smaller than 1 cm. The sequential use of composts suggests that bioaugmentation played a key role in enhancing degradation, as the second compost was inoculated with mature compost from the first trial. Metagenomic analysis identified bacterial genera such as Sinibacillus, Desulfotomaculum, Bdellovibrio, and Proteiniborus, as well as thermotolerant fungi such as Thermomyces, all of which are associated with the enzymatic degradation of proteinaceous materials. In addition, a collection of cultivable strains was obtained, including Bacillus pumilus and Rhodococcus rhodochrous, known for their resistance to heavy metals such as chromium, as well as others like Pseudomonas aeruginosa and Acinetobacter, previously described as degraders of complex substrates. Scanning Electron Microscopy (SEM) images revealed extensive colonisation of leather surfaces by bacteria forming dense biofilms and producing extracellular polymeric substances. These observations confirm that microbial activity was directly involved in the degradation of the leather matrix.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org: Figure S1. Temperature profile of the 2 L Dewar composting system over 1056 hours. Thermophilic conditions (≥50 °C) were rapidly achieved but exhibited frequent fluctuations. Reinoculations (n = 5) were performed when temperature dropped below 50 °C, approximately at hours 160, 288, 576, 720 and 864. Figure S2. Temperature profile of the 40 L Dewar composting system over 1056 hours. Sustained thermophilic conditions were observed, with peak temperatures exceeding 70 °C. Reinoculations (n = 2) were carried out when temperature dropped below 55 °C, at approximately hours 384 and 672. Figure S3. Rarefaction curves based on OTU richness for all compost samples. Curves are shown as interpolated (solid lines) and extrapolated (dashed lines), indicating sequencing depth was sufficient to capture the microbial diversity of each sample. Sample identifiers (ine17–ine26) correspond to samples from 2 L and 40 L Dewar vessels (Table 2). Figure S4. Alpha diversity metrics (Observed richness, Shannon index, Simpson index) at ASV level for 2 L Dewar vessel samples (ine17–ine20) grouped by sample type. Control compost samples show higher diversity compared to leather compost samples. Figure S5. Alpha diversity metrics (Observed richness, Shannon index, Simpson index) at ASV level for 40 L Dewar vessel samples (ine21–ine26) grouped by sample type. Control compost samples show higher diversity compared to leather compost samples. Figure S6. Beta diversity analysis based on PCoA (Principal Coordinates Analysis) of Bray–Curtis dissimilarity for samples ine17–ine20 (2 L Dewar vessel). Control and leather compost samples from two distinct clusters, indicating compositional differences in microbial communities. Figure S7. Beta diversity analysis based on PCoA (Principal Coordinates Analysis) of Bray–Curtis dissimilarity for samples ine21–ine26 (40 L Dewar vessel). Control and leather compost samples cluster separately, indicating consistent differences in microbial community composition across treatments. Figure S8. Relative abundances of bacterial phyla in samples ine17–ine20 (2 L Dewar vessel). Values are expressed as proportions of total ASVs per sample. Figure S9. Relative abundances of bacterial genera in samples ine17–ine20 (2 L Dewar vessel). “Uncultured” category aggregates all unclassified or uncultured ASVs at the genus level. Figure S10. Relative abundances of bacterial phyla in samples ine21–ine26 (40 L Dewar vessel). Values are expressed as proportions of total ASVs per sample. Figure S11. Relative abundances of bacterial genera in samples ine21–ine26 (40 L Dewar vessel). “Uncultured” category aggregates all unclassified or uncultured ASVs at the genus level. Figure S12. Rarefaction curves at the OTU level for fungal samples ine17–ine26. Solid lines indicate interpolated richness; dashed lines indicate extrapolated richness. All curves reached saturation, confirming adequate sequencing depth. Sample identifiers (ine17–ine26) correspond to samples from 2 L and 40 L Dewar vessels (Table 2). Figure S13. Alpha diversity (Observed richness, Shannon index, Simpson index) at ASV level for fungal samples ine17–ine20 (2 L Dewar vessel). Figure S14. Alpha diversity (Observed richness, Shannon index, Simpson index) at ASV level for fungal samples ine21–ine26 (40 L Dewar vessel). Figure S15. Beta diversity analysis based on Bray–Curtis dissimilarity for fungal samples ine17–ine20 (2 L Dewar vessel). PCoA shows clear clustering by sample type. Figure S16. Beta diversity analysis based on Bray–Curtis dissimilarity for fungal samples ine21–ine26 (40 L Dewar vessel). PCoA shows distinct clustering between control compost and leather compost samples. Figure S17. Relative abundances of fungal phyla in samples ine17–ine20 (2 L Dewar vessel). Expressed as percentage of total ASVs per sample. Figure S18. Relative abundances of fungal genera in samples ine17–ine20 (2 L Dewar vessel). The “uncultured” category includes unclassified ASVs at the genus level. Figure S19. Relative abundances of fungal phyla in samples ine21–ine26 (40 L Dewar vessel). Expressed as percentage of total ASVs per sample. Figure S20. Relative abundances of fungal genera in samples ine21–ine26 (40 L Dewar vessel). The “uncultured” category includes unclassified ASVs at the genus level. Table S1. Dimensions and weights of leather sample pieces used in small-scale and large-scale composting experiments
Author Contributions
M.B.-E.: Methodology, investigation, formal analysis, validation, data curation, writing—original draft preparation. I.L.-M.: Methodology, data curation. M.C.: Writing—review and editing; resources. E.O.-C.: Supervision, project administration. F.A.-A.: Supervision, project administration. M.B.: Conceptualization, supervision, writing—review and editing. M.-J.B.: Conceptualization, supervision, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-financed by the European Union through the European Regional Development Fund as part of the Operational Programme of the Valencian Community 2014–2020 and the BIOREQ project, grant number IMDEEA/2021/11, and the University of Alicante, Project UAIND21-02B.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and analysed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Verma, S.K.; Sharma, P.C. Current trends in solid tannery waste management. Crit. Rev. Biotechnol. 2023, 43, 805–822. [Google Scholar] [CrossRef]
- Wrzesińska-Jędrusiak, E.; Czarnecki, M.; Kazimierski, P.; Bandrów, P.; Szufa, S. The Circular Economy in the Management of Waste from Leather Processing. Energies 2023, 16, 564. [Google Scholar] [CrossRef]
- C Chojnacka, K.; Skrzypczak, D.; Mikula, K.; Witek-Krowiak, A.; Izydorczyk, G.; Kuligowski, K.; Bandrów, P.; Kułażyński, M. Progress in sustainable technologies of leather wastes valorization as solutions for the circular economy. J. Clean. Prod. 2021, 313, 127902. [Google Scholar] [CrossRef]
- Thomasset, A.; Benayoun, S. Review: Leather sustainability, an industrial ecology in process. J. Ind. Ecol. 2024, 28, 1842–1856. [Google Scholar] [CrossRef]
- Abo Elazm, A.; Zaki, S.; Abd El-Rahim, W.M.; Rostom, M.; Sabbor, A.T.; Moawad, H.; Sedik, M.Z. Bioremediation of Hexavalent Chromium Widely Discharged in Leather Tanning Effluents. Egypt. J. Chem. 2020, 63, 2201–2212. [Google Scholar] [CrossRef]
- Bakshi, A.; Panigrahi, A.K. Chromium Contamination in Soil and Its Bioremediation: An Overview. In Advances in Bioremediation and Phytoremediation for Sustainable Soil Management; Malik, J.A., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 229–248. ISBN 978-3-030-89984-4. [Google Scholar]
- Biškauskaitė, R.; Valeika, V. Wet Blue Enzymatic Treatment and Its Effect on Leather Properties and Post-Tanning Processes. Materials 2023, 16, 2301. [Google Scholar] [CrossRef] [PubMed]
- Ambone, T.; Joseph, S.; Deenadayalan, E.; Mishra, S.; Jaisankar, S.; Saravanan, P. Polylactic Acid (PLA) Biocomposites Filled with Waste Leather Buff (WLB). J. Polym. Environ. 2017, 25, 1099–1109. [Google Scholar] [CrossRef]
- Santos, R.J.; Agostini, D.L.S.; Cabrera, F.C.; Budemberg, E.R.; Job, A.E. Recycling Leather Waste: Preparing and Studying on the Microstructure, Mechanical, and Rheological Properties of Leather Waste/Rubber Composite. Polym. Compos. 2015, 36, 2275–2281. [Google Scholar] [CrossRef]
- Ayele, M.; Limeneh, D.Y.; Tesfaye, T.; Mengie, W.; Abuhay, A.; Haile, A.; Gebino, G. A Review on Utilization Routes of the Leather Industry Biomass. Adv. mater. sci. eng. 2021, 1503524. [Google Scholar] [CrossRef]
- Rigueto, C.V.T.; Rosseto, M.; Krein, D.D.C.; Ostwald, B.E.P.; Massuda, L.A.; Zanella, B.B.; Dettmer, A. Alternative uses for tannery wastes: a review of environmental, sustainability, and science. J. Leather Sci. Eng. 2020, 2, 21. [Google Scholar] [CrossRef]
- Moktadir, Md.A.; Ahmadi, H.B.; Sultana, R.; Zohra, F.-T.-; Liou, J.J.H.; Rezaei, J. Circular economy practices in the leather industry: A practical step towards sustainable development. J. Clean. Prod. 2020, 251, 119737. [Google Scholar] [CrossRef]
- Hansen, É.; de Aquim, P.M.; Gutterres, M. Environmental assessment of water, chemicals and effluents in leather post-tanning process: A review. Environ. Impact Assess. Rev. 2021, 89, 106597. [Google Scholar] [CrossRef]
- Sokač, T.; Valinger, D.; Benković, M.; Jurina, T.; Gajdoš Kljusurić, J.; Radojčić Redovniković, I.; Jurinjak Tušek, A. Application of Optimization and Modeling for the Composting Process Enhancement. Processes 2022, 10, 229. [Google Scholar] [CrossRef]
- Goswami, L.; Mukhopadhyay, R.; Bhattacharya, S.S.; Das, P.; Goswami, R. Detoxification of chromium-rich tannery industry sludge by Eudrillus eugeniae: Insight on compost quality fortification and microbial enrichment. Bioresour. Technol. 2018, 266, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Vico, A.; Maestre-Lopez, M.I.; Arán-Ais, F.; Orgilés-Calpena, E.; Bertazzo, M.; Marhuenda-Egea, F.C. Assessment of the Biodegradability and Compostability of Finished Leathers: Analysis Using Spectroscopy and Thermal Methods. Polymers 2024, 16, 1908. [Google Scholar] [CrossRef]
- Alugoju, P.; Rao, C.S.V.; Babu, R.; Thankappan, R. Assessment of Biodegradability of Synthetic Tanning Agents Used in Leather Tanning Process. International Journal of Engineering and Technology 2011, 3, 302–308. [Google Scholar]
- Chen, X.; Zhao, Y.; Zhang, C.; Zhang, D.; Yao, C.; Meng, Q.; Zhao, R.; Wei, Z. Speciation, toxicity mechanism and remediation ways of heavy metals during composting: A novel theoretical microbial remediation method is proposed. J. Environ. Manage. 2020, 272, 111109. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Liu, X.; Wang, Y. Amphoteric functional polymers for leather wet finishing auxiliaries: A review. Polym. Adv. Technol. 2021, 32, 1951–1964. [Google Scholar] [CrossRef]
- Wieczorek, D.; Gendaszewska, D.; Miśkiewicz, K.; Słubik, A.; Ławińska, K. Biotransformation of Protein-Rich Waste by Yarrowia Lipolytica IPS21 to High-Value Products—Amino Acid Supernatants. Microbiol. Spectr. 2023, 11, e02749–23. [Google Scholar] [CrossRef] [PubMed]
- Sardroudi, N.P.; Sorolla, S.; Casas, C.; Bacardit, A. A Study of the Composting Capacity of Different Kinds of Leathers, Leatherette and Alternative Materials. Sustainability 2024, 16, 2324. [Google Scholar] [CrossRef]
- Rosu, L.; Varganici, C.-D.; Crudu, A.-M.; Rosu, D. Influence of different tanning agents on bovine leather thermal degradation. J. Therm. Anal. Calorim. 2018, 134, 583–594. [Google Scholar] [CrossRef]
- Codreanu (Manea), A.-M.N.; Stefan, D.S.; Kim, L.; Stefan, M. Depollution of Polymeric Leather Waste by Applying the Most Current Methods of Chromium Extraction. Polymers 2024, 16, 1546. [Google Scholar] [CrossRef]
- Piccin, J.S.; Feris, L.A.; Cooper, M.; Gutterres, M. Dye Adsorption by Leather Waste: Mechanism Diffusion, Nature Studies, and Thermodynamic Data. J. Chem. Eng. Data 2013, 58, 873–882. [Google Scholar] [CrossRef]
- Halldórsdóttir, H.H.; Williams, R.; Greene, E.M.; Taylor, G. Rapid deterioration in buried leather: archaeological implications. RSC Adv. 2024, 14, 3762–3770. [Google Scholar] [CrossRef]
- Akhtar, M.A.; Butt, M.Q.S.; Afroz, A.; Rasul, F.; Irfan, M.; Sajjad, M.; Zeeshan, N. Approach towards sustainable leather: Characterization and effective industrial application of proteases from Bacillus sps. for ecofriendly dehairing of leather hide. Int. J. Biol. Macromol. 2024, 266, 131154. [Google Scholar] [CrossRef]
- Alibardi, L.; Cossu, R. Pre-treatment of tannery sludge for sustainable landfilling. Waste Manag. 2016, 52, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Kanagaraj, J.; Panda, R.C.; Kumar, M.V. Trends and advancements in sustainable leather processing: Future directions and challenges—A review. J. Environ. Chem. Eng. 2020, 8, 104379. [Google Scholar] [CrossRef]
- Blair, E.M.; Dickson, K.L.; O’Malley, M.A. Microbial communities and their enzymes facilitate degradation of recalcitrant polymers in anaerobic digestion. Curr. Opin. Microbiol. 2021, 64, 100–108. [Google Scholar] [CrossRef]
- Biyada, S.; Merzouki, M.; Dėmčėnko, T.; Vasiliauskienė, D.; Ivanec-Goranina, R.; Urbonavičius, J.; Marčiulaitienė, E.; Vasarevičius, S.; Benlemlih, M. Microbial community dynamics in the mesophilic and thermophilic phases of textile waste composting identified through next-generation sequencing. Sci. Rep. 2021, 11, 23624. [Google Scholar] [CrossRef]
- Finore, I.; Feola, A.; Russo, L.; Cattaneo, A.; Di Donato, P.; Nicolaus, B.; Poli, A.; Romano, I. Thermophilic bacteria and their thermozymes in composting processes: a review. Chem. Biol. Technol. Agric. 2023, 10, 7. [Google Scholar] [CrossRef]
- Steiniger, B.; Hupfauf, S.; Insam, H.; Schaum, C. Exploring Anaerobic Digestion from Mesophilic to Thermophilic Temperatures—Operational and Microbial Aspects. Fermentation 2023, 9, 798. [Google Scholar] [CrossRef]
- Nnaji, N.D.; Anyanwu, C.U.; Miri, T.; Onyeaka, H. Mechanisms of Heavy Metal Tolerance in Bacteria: A Review. Sustainability 2024, 16, 11124. [Google Scholar] [CrossRef]
- Azarbad, H.; van Gestel, C.A.M.; Niklińska, M.; Laskowski, R.; Röling, W.F.M.; van Straalen, N.M. Resilience of Soil Microbial Communities to Metals and Additional Stressors: DNA-Based Approaches for Assessing “Stress-on-Stress” Responses. Int. J. Mol. Sci. 2016, 17, 933. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Paul, A.K. Microbial extracellular polymeric substances: central elements in heavy metal bioremediation. Indian J. Microbiol. 2008, 48, 49–64. [Google Scholar] [CrossRef]
- Kang, D.; Jacquiod, S.; Herschend, J.; Wei, S.; Nesme, J.; Sørensen, S.J. Construction of Simplified Microbial Consortia to Degrade Recalcitrant Materials Based on Enrichment and Dilution-to-Extinction Cultures. Front. Microbiol. 2020, 10, 3010. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, J.; Achinas, S.; Zhang, Z.; Krooneman, J.; Euverink, G.J.W. The biomethanation of cow manure in a continuous anaerobic digester can be boosted via a bioaugmentation culture containing Bathyarchaeota. Sci. Total Environ. 2020, 745, 141042. [Google Scholar] [CrossRef]
- Yang, G.; Chen, J.; Zhou, S. Novibacillus thermophilus gen. nov., sp. nov., a Gram-staining-negative and moderately thermophilic member of the family Thermoactinomycetaceae. Int. J. Syst. Evol. Microbiol. 2015, 65, 2591–2597. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Lin, J.; Wang, W.; Huang, H.; Li, S. Biodegradation of aliphatic and polycyclic aromatic hydrocarbons by the thermophilic bioemulsifier-producing Aeribacillus pallidus strain SL-1. Ecotoxicol. Environ. Saf. 2020, 189, 109994. [Google Scholar] [CrossRef] [PubMed]
- Gomez del Pulgar, E.M.; Saadeddin, A. The cellulolytic system of Thermobifida fusca. Crit. Rev. Microbiol. 2014, 40, 236–247. [Google Scholar] [CrossRef] [PubMed]
- McFall, A.; Coughlin, S.A.; Hardiman, G.; Megaw, J. Strategies for biofilm optimization of plastic-degrading microorganisms and isolating biofilm formers from plastic-contaminated environments. Sustain. Microbiol. 2024, 1, qvae012. [Google Scholar] [CrossRef]
- Hassan, M.A.; Haroun, B.M.; Amara, A.A.; Serour, E.A. Production and Characterization of Keratinolytic Protease from New Wool-Degrading Bacillus Species Isolated from Egyptian Ecosystem. BioMed Res. Int. 2013, 2013, 175012. [Google Scholar] [CrossRef]
- Akram, F.; Haq, I. ul; Hayat, A.K.; Ahmed, Z.; Jabbar, Z.; Baig, I.M.; Akram, R. Keratinolytic Enzyme from a Thermotolerant Isolate Bacillus sp. NDS-10: An Efficient Green Biocatalyst for Poultry Waste Management, Laundry and Hide-dehairing Applications. Waste Biomass Valor 2021, 12, 5001–5018. [Google Scholar] [CrossRef]
- McPhillips, K.; Waters, D.M.; Parlet, C.; Walsh, D.J.; Arendt, E.K.; Murray, P.G. Purification and Characterisation of a β-1,4-Xylanase from Remersonia Thermophila CBS 540.69 and Its Application in Bread Making. Appl. Biochem. Biotechnol. 2014, 172, 1747–1762. [Google Scholar] [CrossRef] [PubMed]
- van den Brink, J.; Facun, K.; de Vries, M.; Stielow, J.B. Thermophilic growth and enzymatic thermostability are polyphyletic traits within Chaetomiaceae. Fungal Biol. 2015, 119, 1255–1266. [Google Scholar] [CrossRef]
- Kong, Z.; Wang, X.; Wang, M.; Chai, L.; Wang, X.; Liu, D.; Shen, Q. Bacterial ecosystem functioning in organic matter biodegradation of different composting at the thermophilic phase. Bioresour. Technol. 2020, 317, 123990. [Google Scholar] [CrossRef]
- Radhika, V.; Subramanian, S.; Natarajan, K.A. Bioremediation of zinc using Desulfotomaculum nigrificans: Bioprecipitation and characterization studies. Water Res. 2006, 40, 3628–3636. [Google Scholar] [CrossRef]
- Patel, A.B.; Singh, S.; Patel, A.; Jain, K.; Amin, S.; Madamwar, D. Synergistic biodegradation of phenanthrene and fluoranthene by mixed bacterial cultures. Bioresour. Technol. 2019, 284, 115–120. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, D.; Tang, W.; Wang, L.; Li, Q.; Lu, Z.; Liu, H.; Zhong, Y.; He, T.; Guo, S. Phytoremediation of cadmium-polluted soil assisted by D-gluconate-enhanced Enterobacter cloacae colonization in the Solanum nigrum L. rhizosphere. Sci. Total Environ. 2020, 732, 139265. [Google Scholar] [CrossRef]
- Kiiskinen, L.-L.; Palonen, H.; Linder, M.; Viikari, L.; Kruus, K. Laccase from Melanocarpus albomyces binds effectively to cellulose. FEBS Lett. 2004, 576, 251–255. [Google Scholar] [CrossRef]
- Dobrowolski, R.; Szcześ, A.; Czemierska, M.; Jarosz-Wikołazka, A. Studies of cadmium(II), lead(II), nickel(II), cobalt(II) and chromium(VI) sorption on extracellular polymeric substances produced by Rhodococcus opacus and Rhodococcus rhodochrous. Bioresour. Technol. 2017, 225, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Efe, D. Potential Plant Growth-Promoting Bacteria with Heavy Metal Resistance. Curr. Microbiol. 2020, 77, 3861–3868. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Merwad, A.-R.M.; Semida, W.M.; Ibrahim, S.A.; El-Saadony, M.T.; Rady, M.M. Heavy metals-resistant bacteria (HM-RB): Potential bioremediators of heavy metals-stressed Spinacia oleracea plant. Ecotoxicol. Environ. Saf. 2020, 198, 110685. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.; Hodgson, D.J.; Buckling, A. Social evolution of toxic metal bioremediation in Pseudomonas aeruginosa. Proc. R. Soc. B 2014, 281, 20140858. [Google Scholar] [CrossRef]
- Sun, L.; Zheng, Y.; Yu, X. Flocculation-bio-treatment of heavy metals-vacuum preloading of the river sediments. Ecotoxicol. Environ. Saf. 2020, 201, 110810. [Google Scholar] [CrossRef]
- Pang, B.; Lv, L.; Pang, C.; Ye, F.; Shang, C. Optimization of Growth Conditions of Acinetobacter Sp. Cr1 for Removal of Heavy Metal Cr Using Central Composite Design. Curr Microbiol 2021, 78, 316–322. [Google Scholar] [CrossRef]
- Wu, P.; Wang, Z.; Bhatnagar, A.; Jeyakumar, P.; Wang, H.; Wang, Y.; Li, X. Microorganisms-Carbonaceous Materials Immobilized Complexes: Synthesis, Adaptability and Environmental Applications. J. Hazard. Mater. 2021, 416, 125915. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Gupta, A. Evaluation of Acinetobacter Sp. B9 for Cr (VI) Resistance and Detoxification with Potential Application in Bioremediation of Heavy-Metals-Rich Industrial Wastewater. Environ Sci Pollut Res 2013, 20, 6628–6637. [Google Scholar] [CrossRef] [PubMed]
- Che Hussian, C.H.A.; Leong, W.Y. Thermostable enzyme research advances: a bibliometric analysis. J. Genet. Eng. Biotechnol. 2023, 21, 37. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Shukla, P. A comparative analysis of heavy metal bioaccumulation and functional gene annotation towards multiple metal resistant potential by Ochrobactrum intermedium BPS-20 and Ochrobactrum ciceri BPS-26. Bioresour. Technol. 2021, 320, 124330. [Google Scholar] [CrossRef]
- Kumar, M.; Revathi, K.; Khanna, S. Biodegradation of cellulosic and lignocellulosic waste by Pseudoxanthomonas sp R-28. Carbohydr. Polym. 2015, 134, 761–766. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Cheng, Y.-C.; Yang, W.-L.; Wang, M.-Y. Surface Defect Detection of Wet-Blue Leather Using Hyperspectral Imaging. IEEE Access 2021, 9, 127685–127702. [Google Scholar] [CrossRef]
- Mehta, M.; Naffa, R.; Maidment, C.; Holmes, G.; Waterland, M. Raman and Atr-Ftir Spectroscopy towards Classification of Wet Blue Bovine Leather Using Ratiometric and Chemometric Analysis. J. Leather Sci. Eng. 2020, 2, 3. [Google Scholar] [CrossRef]
- NZYSoil gDNA Isolation Kit. Available online: https://www.nzytech.com/en/mb21802-nzy-soil-gdna-isolation-kit/ (accessed on 13 June 2024).
- Church, D.L.; Cerutti, L.; Gürtler, A.; Griener, T.; Zelazny, A.; Emler, S. Performance and Application of 16S rRNA Gene Cycle Sequencing for Routine Identification of Bacteria in the Clinical Microbiology Laboratory. Clin. Microbiol. Rev. 2020, 33, e00053–19. [Google Scholar] [CrossRef] [PubMed]
- Sykes, J.E.; Rankin, S.C. Chapter 4 - Isolation and Identification of Fungi. In Canine and Feline Infectious Diseases, 1st ed.; Sykes, J.E., Ed.; W.B. Saunders: Saint Louis, 2014; pp. 29–36. ISBN 978-1-4377-0795-3. [Google Scholar]
- Satari, L.; Guillén, A.; Vidal-Verdú, À.; Porcar, M. The wasted chewing gum bacteriome. Sci. Rep. 2020, 10, 16846. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS One 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P. vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Preprint at 2022, 3–1.
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Hardersen, S.; La Porta, G. Never underestimate biodiversity: how undersampling affects Bray–Curtis similarity estimates and a possible countermeasure. Eur. Zool. J. 2023, 90, 660–672. [Google Scholar] [CrossRef]
- Garrido-Martín, D.; Calvo, M.; Reverter, F.; Guigó, R. A fast non-parametric test of association for multiple traits. Genome Biol. 2023, 24, 230. [Google Scholar] [CrossRef]
- Nucleotide BLAST: Search Nucleotide Databases Using a Nucleotide Query. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&BLAST_SPEC=GeoBlast&PAGE_TYPE=BlastSearch (accessed on 10 February 2025).
- Vyskočilová, G.; Carşote, C.; Ševčík, R.; Badea, E. Burial-induced deterioration in leather: A FTIR-ATR, DSC, TG/DTG, MHT and SEM study. Herit. Sci. 2022, 10, 7. [Google Scholar] [CrossRef]
Figure 1.
Taxonomic distribution at the phylum level for samples ine17–ine20 (small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
Figure 1.
Taxonomic distribution at the phylum level for samples ine17–ine20 (small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
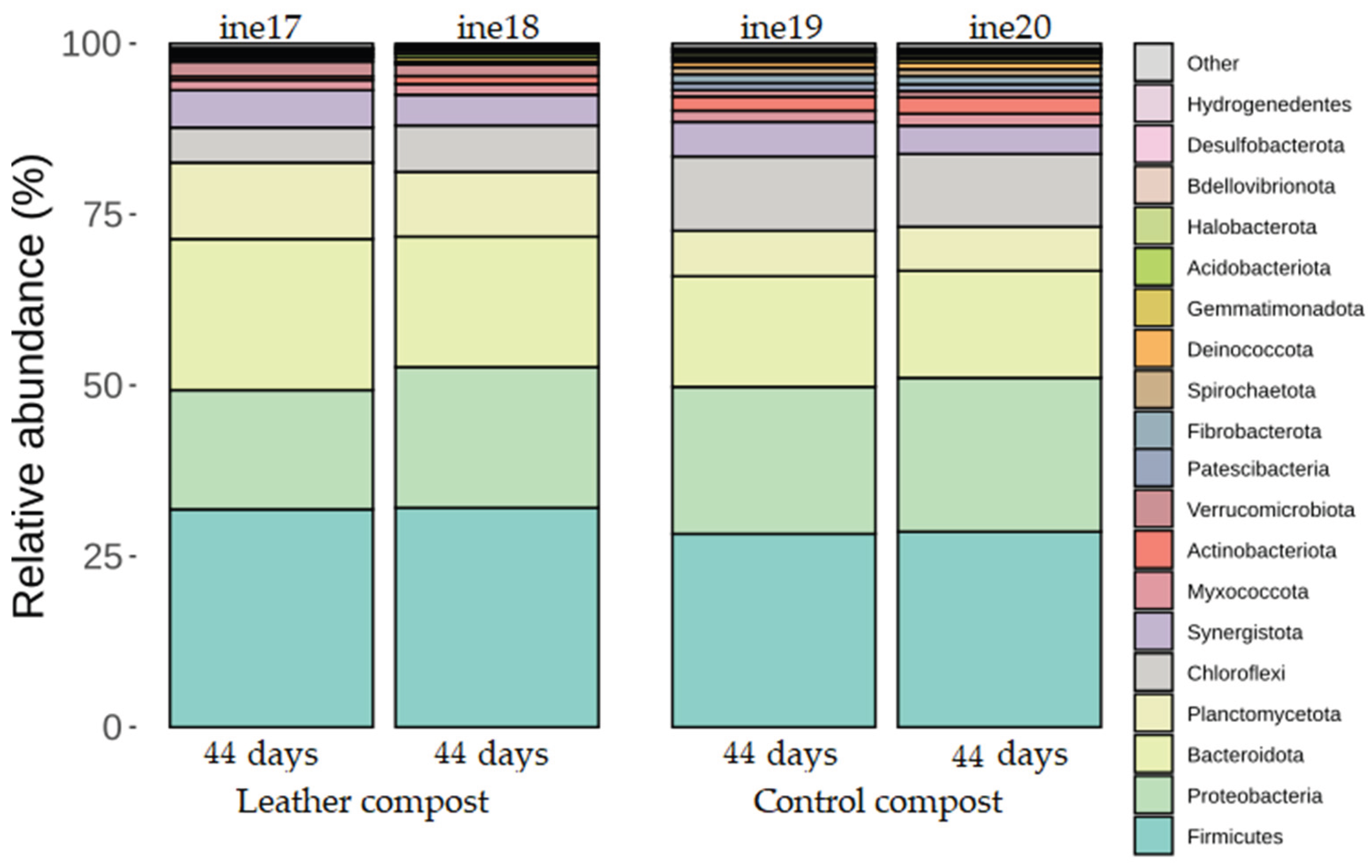
Figure 2.
Taxonomic distribution at the genus level for samples ine17–ine20 (small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
Figure 2.
Taxonomic distribution at the genus level for samples ine17–ine20 (small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
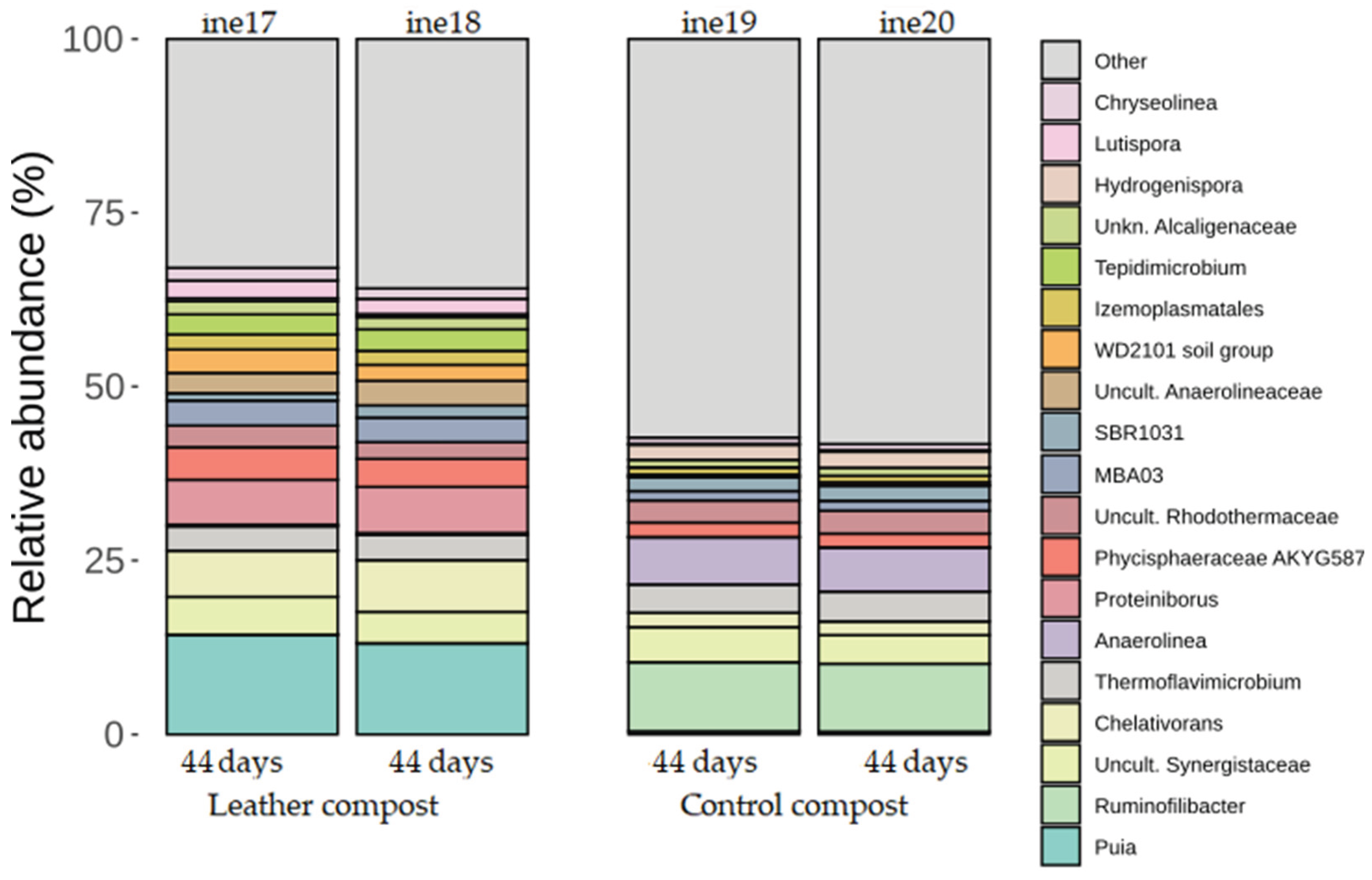
Figure 3.
Taxonomic distribution at the phylum level for samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
Figure 3.
Taxonomic distribution at the phylum level for samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
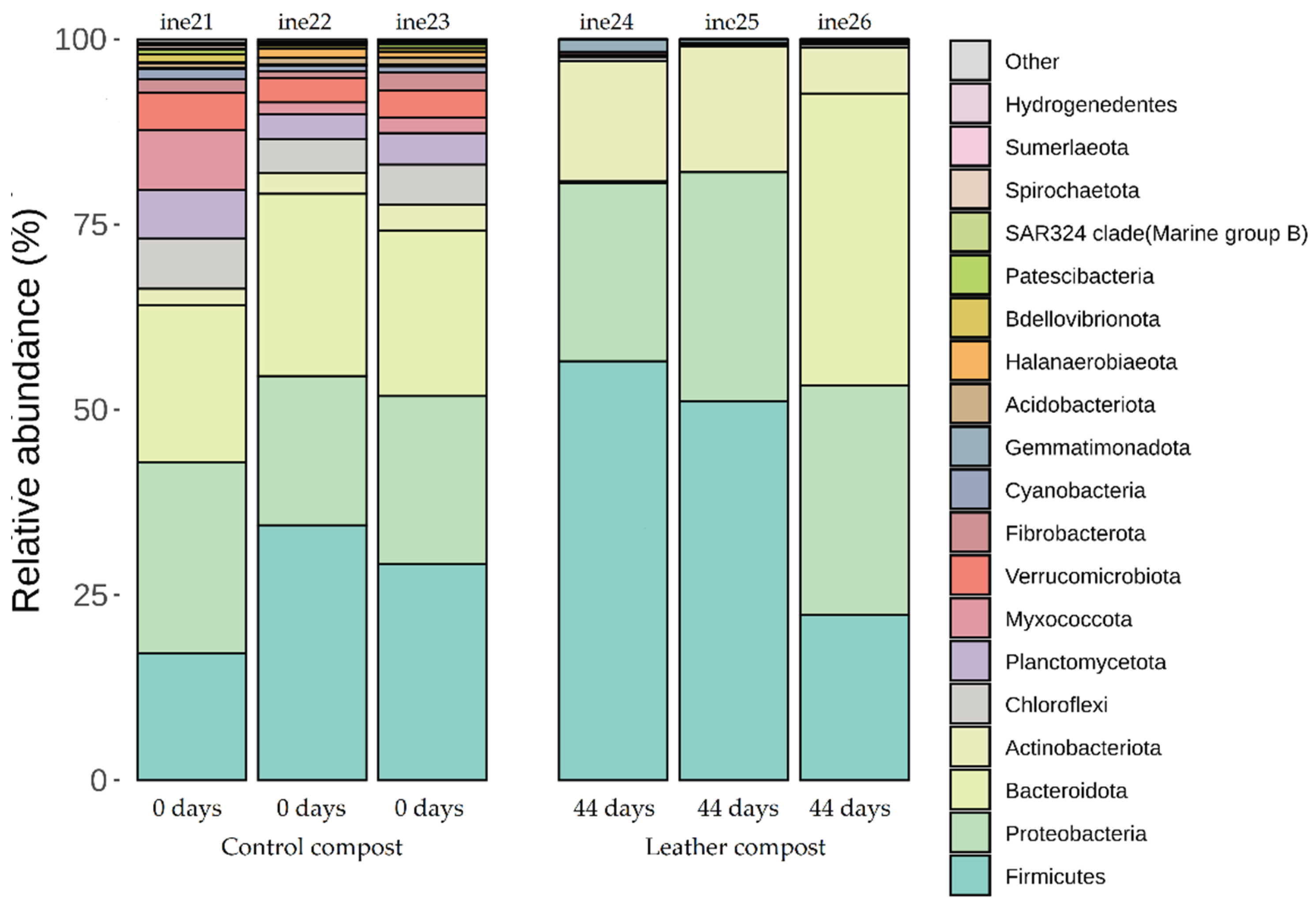
Figure 4.
Taxonomic distribution at the genus level for samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
Figure 4.
Taxonomic distribution at the genus level for samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 16S rRNA gene-based taxonomic profiling. The “Others” category includes bacterial taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
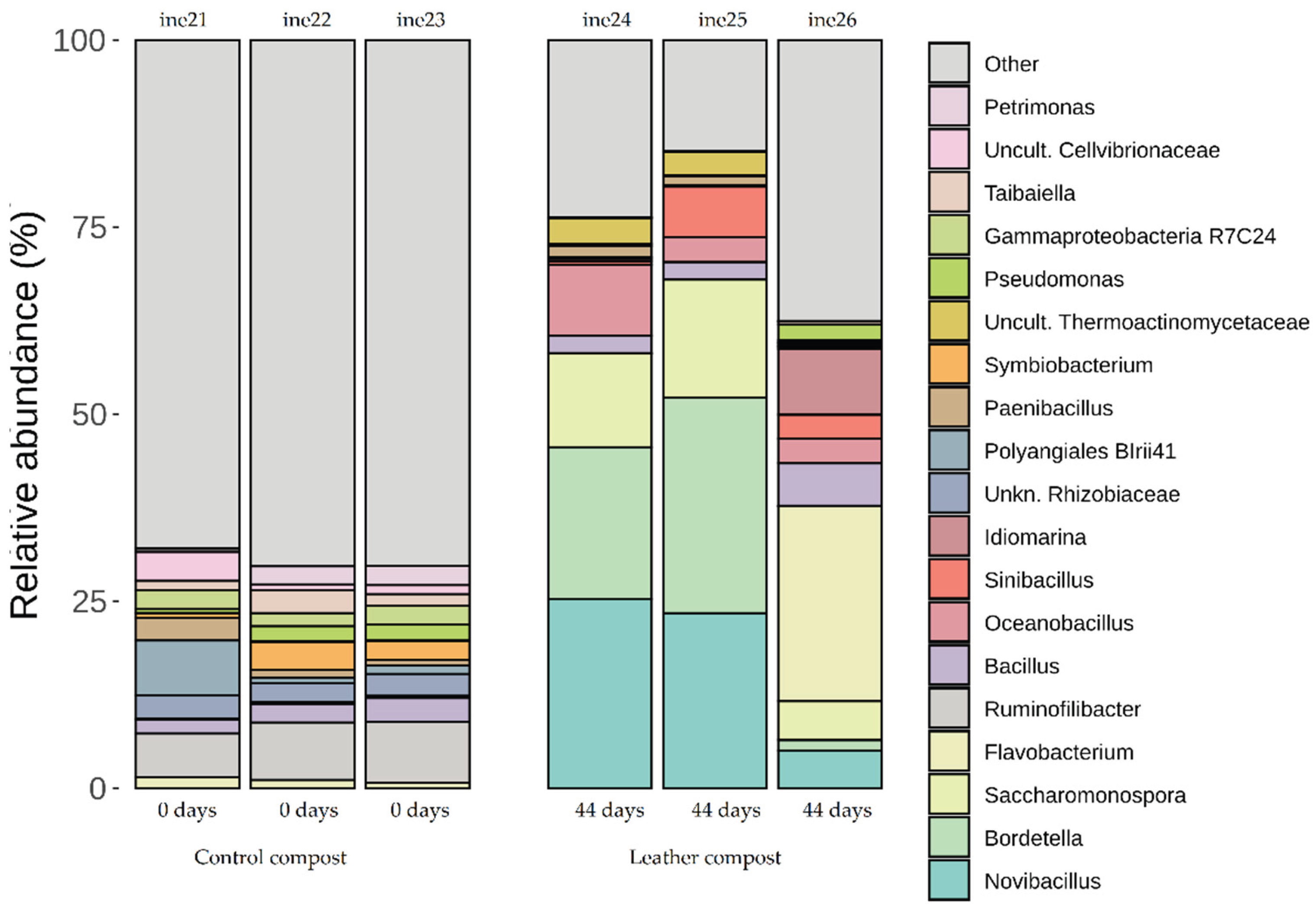
Figure 5.
Taxonomic distribution at the phylum level for samples ine17–ine20 (small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling.
Figure 5.
Taxonomic distribution at the phylum level for samples ine17–ine20 (small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling.
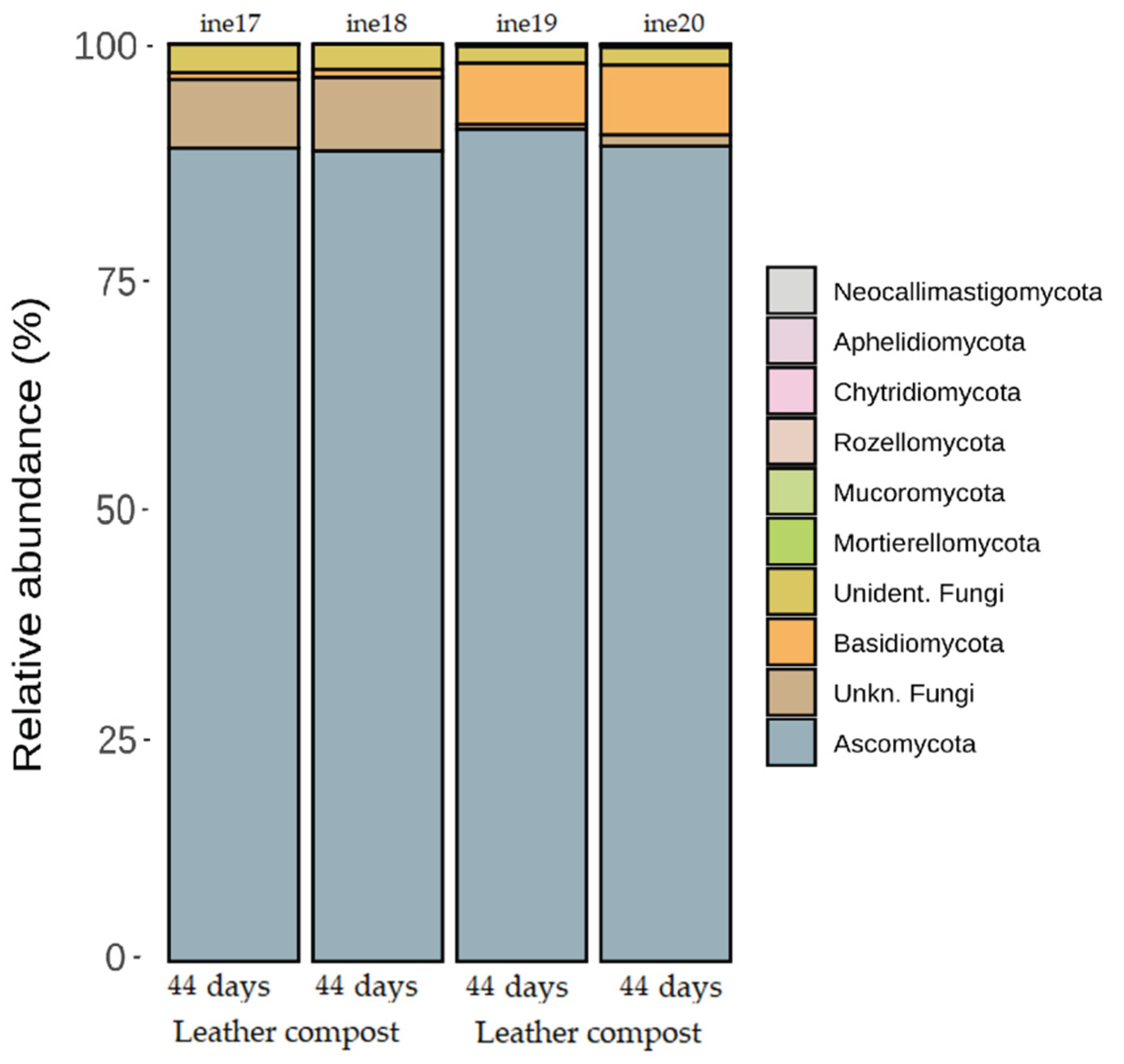
Figure 6.
Taxonomic distribution at the genus level for fungal samples ine17–ine20(small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling. The “Others” category includes fungal taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
Figure 6.
Taxonomic distribution at the genus level for fungal samples ine17–ine20(small-scale 2 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling. The “Others” category includes fungal taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
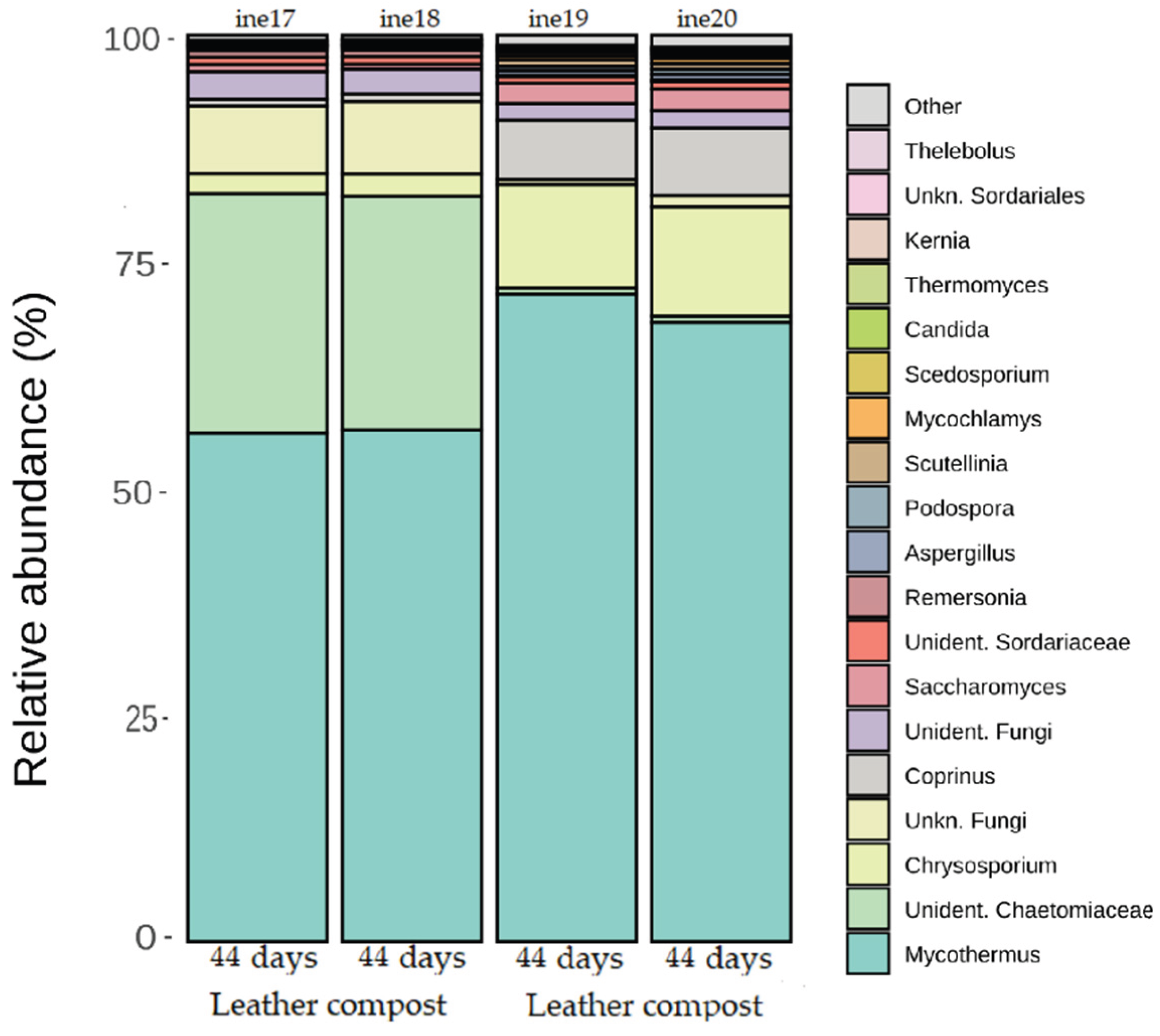
Figure 7.
Taxonomic distribution at the phylum level for fungal samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling.
Figure 7.
Taxonomic distribution at the phylum level for fungal samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling.
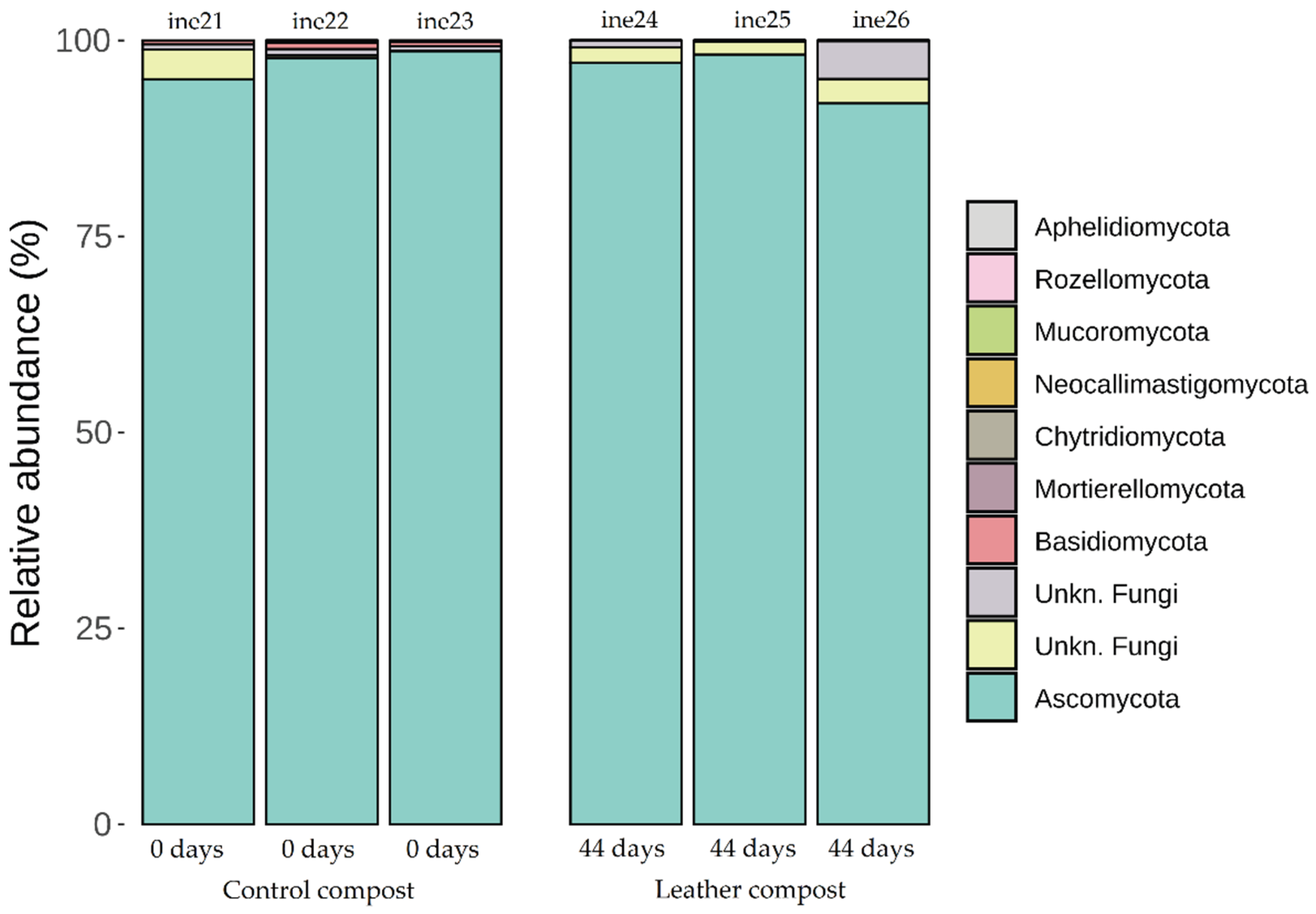
Figure 8.
Taxonomic distribution at the genus level for fungal samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling. The “Others” category includes fungal taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
Figure 8.
Taxonomic distribution at the genus level for fungal samples ine21–ine26 (large-scale 40 L Dewar vessel). All samples shown refer to those analysed for 23S rRNA gene-based taxonomic profiling. The “Others” category includes fungal taxa with annotations that do not reach the phylum level and/or exhibit low relative abundance across samples.
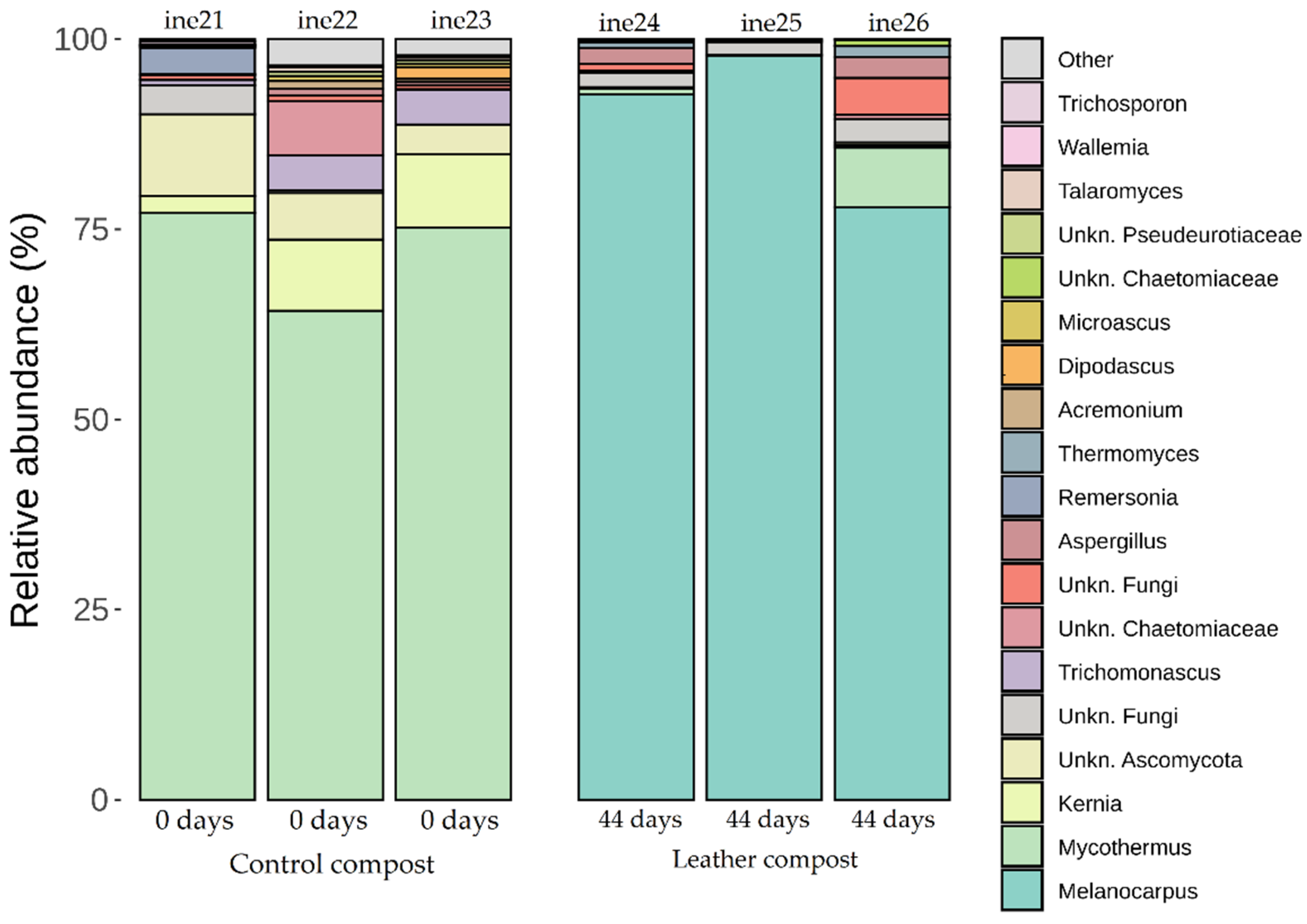
Figure 9.
Leather pieces shown in Figure 13B were composted in a 2 L vessel for 44 days.
Figure 9.
Leather pieces shown in Figure 13B were composted in a 2 L vessel for 44 days.

Figure 10.
Scanning electron microscopy (SEM) images of composted leather fragments. Characteristic collagen fibres of wet blue leather are visible. Images captured at ×2500 magnification.
Figure 10.
Scanning electron microscopy (SEM) images of composted leather fragments. Characteristic collagen fibres of wet blue leather are visible. Images captured at ×2500 magnification.
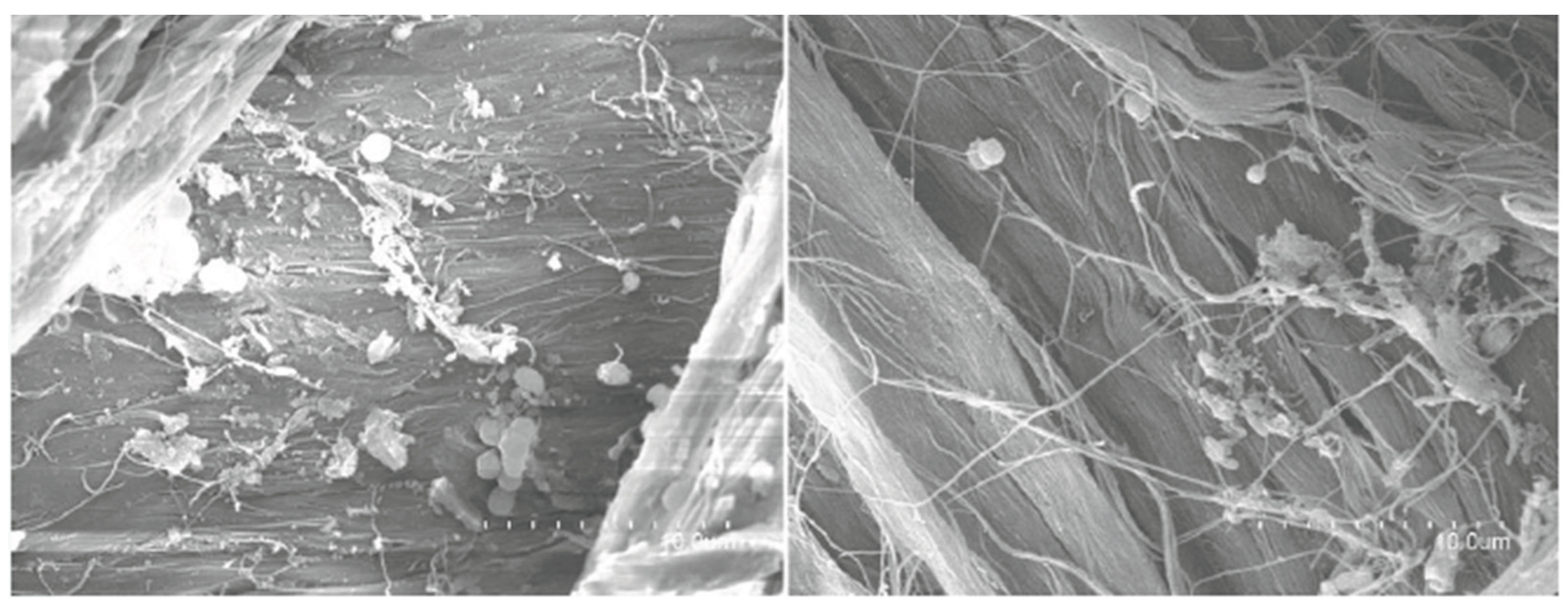
Figure 11.
SEM images of the composted leather surface showing extensive bacterial colonisation and biofilm formation. Images captured at ×3500 magnification.
Figure 11.
SEM images of the composted leather surface showing extensive bacterial colonisation and biofilm formation. Images captured at ×3500 magnification.
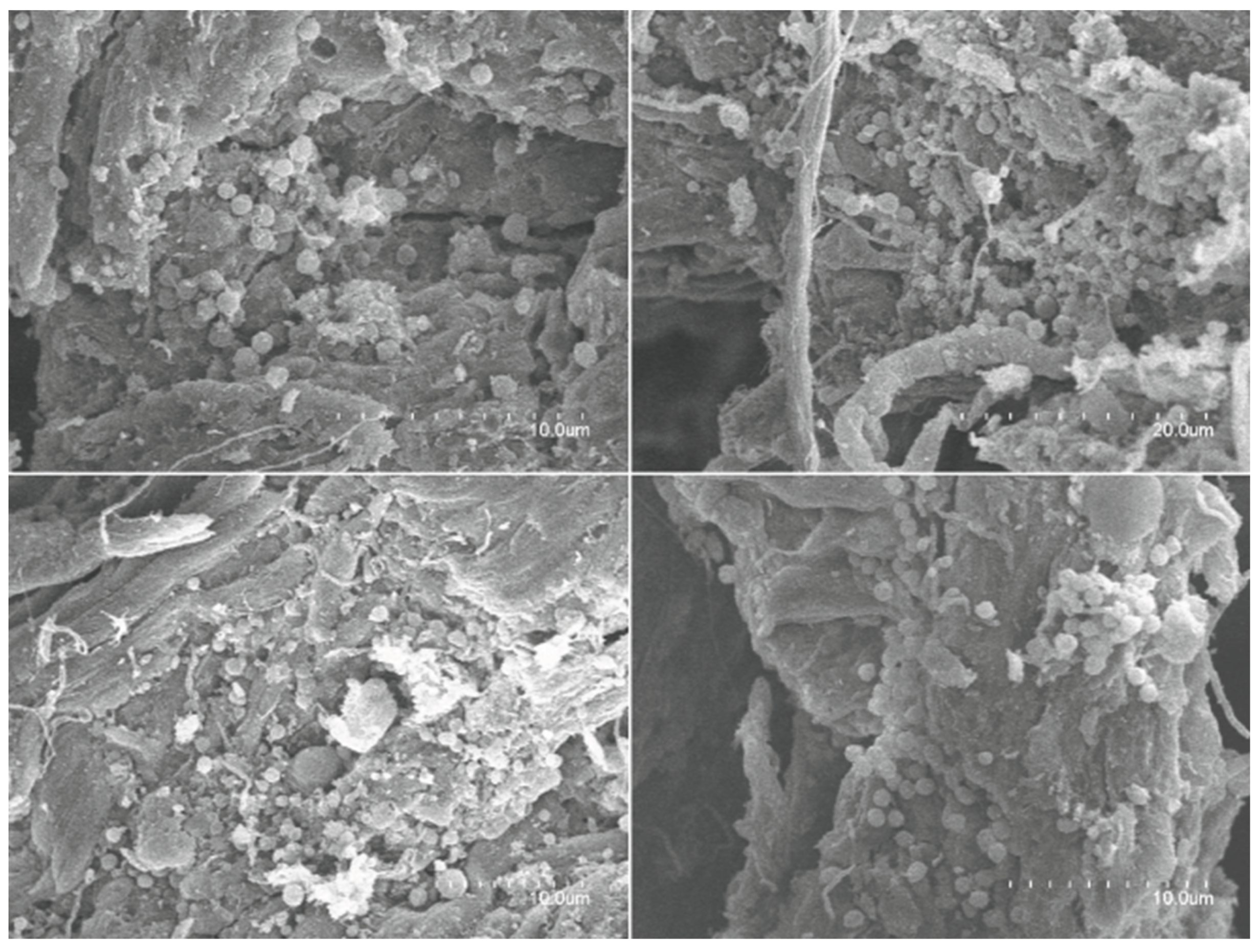
Figure 12.
SEM images of composted leather surface at higher magnification (×7000), revealing dense bacterial coverage and abundant extracellular matrix consistent with mature biofilms.
Figure 12.
SEM images of composted leather surface at higher magnification (×7000), revealing dense bacterial coverage and abundant extracellular matrix consistent with mature biofilms.

Figure 13.
Wet blue leather samples used in composting assays. (A) Three rectangular leather samples (approx. 10 × 5.5 cm) placed in the 40 L Dewar vessel for large-scale composting, with initial weights measured before exposure. (B) One of the three leather strips (7 × 1.5 cm) used in the small-scale composting system (2 L Dewar vessel), fixed vertically on metal rods to ensure stability within the compost matrix.
Figure 13.
Wet blue leather samples used in composting assays. (A) Three rectangular leather samples (approx. 10 × 5.5 cm) placed in the 40 L Dewar vessel for large-scale composting, with initial weights measured before exposure. (B) One of the three leather strips (7 × 1.5 cm) used in the small-scale composting system (2 L Dewar vessel), fixed vertically on metal rods to ensure stability within the compost matrix.


Table 1.
Cultivable strains isolated from degraded leather after composting, from both the 2 L and 40 L compost systems.
Table 1.
Cultivable strains isolated from degraded leather after composting, from both the 2 L and 40 L compost systems.
| Closest Species | Medium | Temp. | Similarity (%) |
|---|---|---|---|
| Rhodococcus rhodochrous, NBRC 16069 | R2A | 30ºC | 98.50 |
| Mammaliicoccus sciuri, DSM 20345 | R2A | 30ºC | 99.84 |
| Bacillus zhangzhouensis, DW5-4 | R2A | 30ºC | 98.87 |
| Pseudomonas aeruginosa, JCM 5962 | R2A | 30ºC | 99.69 |
| Leuconostoc mesenteroides subsp. mesenteroides, ATCC 8293 | R2A | 30ºC | 99.32 |
| Empedobacter brevis, LMG 4011 | TSA | 30ºC | 98.85 |
| Bacillus tequilensis, KCTC 13622 | TSA | 30ºC | 99.53 |
| Acinetobacter beijerinckii, CIP 110307 | TSA | 30ºC | 99.14 |
| Glutamicibacter mishrai S5-52 | TSA | 30ºC | 99.36 |
| Bacillus siamensis KCTC 13613 | TSA | 30ºC | 99.28 |
| Bacillus smithii DSM 4216 | YM | 55ºC | 99.20 |
| Paenibacillus typhae CGMCC 1.11012 | YM | 55ºC | 99.25 |
| Aneurinibacillus thermoaerophilus DSM 10154 | YM | 55ºC | 99.28 |
| Ureibacillus suwonensis DSM 16752 | TSA | 55ºC | 99.09 |
| Chelatococcus composti PC-2 | TSA | 55ºC | 99.66 |
| Caldibacillus hisashii N-11 | GYM | 55ºC | 98.02 |
| Aeribacillus pallidus KCTC 3564 | GYM | 55ºC | 98.83 |
| Chelativorans composti, Nis 3 | GYM | 55ºC | 99.40 |
| Thermomyces lanuginosus (hongo) | YM | 55ºC | 100 |
| Brucella ciceri, Ca-34 | R2A | 30ºC | 97.89 |
| Chryseobacterium bernardetii, NCTC 13530 | R2A | 30ºC | 99.67 |
| Brevibacillus parabrevis, NRRL NRS 605 | R2A | 30ºC | 96.59 |
| Bacillus paralicheniformis, KJ-16 | TSA | 55ºC | 99.10 |
| Aeribacillus composti, N.8 | NAI | 55ºC | 99.25 |
| Bacillus aerius, 24K | YM | 55ºC | 94.03 |
| Weizmannia coagulans, ATCC 7050 | YM | 55ªC | 97.25 |
| Ureibacillus thermosphaericus, DSM 10633 | TSA | 55ªC | 99.81 |
| Aeribacillus pallidus, KCTC 3564 | TSA | 55ªC | 3.4 |
Table 2.
Description of compost samples analysed for DNA extraction and metagenomic sequencing. Samples were collected from two composting systems: a small-scale 2 L Dewar vessel (ine17–ine20) and a large-scale 40 L Dewar vessel (ine21–ine26). Each sample is classified according to its origin (composted leather or control compost) and the duration of composting (0 or 44 days).
Table 2.
Description of compost samples analysed for DNA extraction and metagenomic sequencing. Samples were collected from two composting systems: a small-scale 2 L Dewar vessel (ine17–ine20) and a large-scale 40 L Dewar vessel (ine21–ine26). Each sample is classified according to its origin (composted leather or control compost) and the duration of composting (0 or 44 days).
| SAMPLE NAME | VESSEL | TYPE | TIME (days) |
|---|---|---|---|
| ine17 | 2L | Composted leather | 44 |
| ine18 | 2L | Composted leather | 44 |
| ine19 | 2L | Control compost | 44 |
| ine20 | 2L | Control compost | 44 |
| ine21 | 40L | Control compost | 0 |
| ine22 | 40L | Control compost | 0 |
| ine23 | 40L | Control compost | 0 |
| ine24 | 40L | Composted leather | 44 |
| ine25 | 40L | Composted leather | 44 |
| ine26 | 40L | Composted leather | 44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



