Submitted:
28 August 2025
Posted:
29 August 2025
You are already at the latest version
Abstract
Microalgae are seen as a promising source of sustainable essential lipids for animal and human nutrition. These advantages, however, differ depending on the strain and growing conditions. Furthermore, the link between microalgae lipid profile and their effect on target organisms during dietary consumption is still not well understood. This study examined the lipid content and antioxidant properties of four microalgae strains cultured on 3N-BBM+V medium, and the effect of their dietary inclusions on the lifespan, body mass, and fertility of the common fruit fly Drosophila melanogaster. It was shown that C. vulgaris, N. limnetica, S. communis, T. obliquus exhibit total lipids at 26.7 ± 2.8%, 27.2 ± 0.9%, 16.6 ± 3.0%, and 22.6 ± 0.9% of dry biomass, respectively. All microalgal species produced unsaturated C15:0, C16:0, monosaturated C18:2n6 (omega-6), and C18:1n9 (omega-9) but at different proportions. Feeding D. melanogaster with C. vulgaris, S. communis, or T. obliquus biomass at 3 mg/mL inclusion extended the female median lifespan by 45.9%, 8.1%, and 27.0%, respectively. A more significant extension was observed for fly males, where adding biomass of C. vulgaris, N. limnetica, S. communis, and T. obliquus increased the lifespan of the flies by up to 75.0%, 25%, 29.2% and 50.0% re-spectively. The fertility of flies fed with T. obliquus and S. communis was also increased by 20% and 18% respectively. Meanwhile, no significant difference in the body mass of flies fed with microalgae was detected compared to the control.

Keywords:
microalgae
; lipid concentrations
; fatty acid profile
; Drosophila melanogaster
1. Introduction
Microalgae have attracted increasing attention as a natural source of valuable biomolecules and biomass for human and animal nutrition, as they provide lipids, proteins, polysaccharides, pigments, vitamins, and minerals [1,2]. Oleaginous species are of particular interest because, in addition to other bioactive properties, they contain high levels of essential lipids for dietary purposes [3,4]. Continuous efforts are therefore directed toward identifying such species and characterizing their fatty acid (FA) profiles [5]. For effective screening, however, a rapid and cost-efficient method to evaluate their effects on the whole organism is required. Drosophila melanogaster is increasingly recognized as a model organism in food and nutrition research, owing to its relatively short lifespan and generation time, and genetic homology with humans, which make it particularly suitable for such studies [6,7]. Although widely employed in nutritional research, only a limited number of studies have investigated the impact of microalgal supplementation on D. melanogaster [8,9]. The present study aims to analyze the lipid content and FA profiles of four microalgal strains and to assess their effects on key health parameters of D. melanogaster, including lifespan, body mass, and fecundity. Importantly, this work provides new insights into the link between microalgal lipid composition and organism-level health outcomes, highlighting the potential of D. melanogaster as a rapid model for evaluating lipid sources.
2. Materials and Methods
2.1. Microalgae Species and Growth Conditions
Nannochloropsis limnetica (SAG 18.99) was purchased from the Culture Collection of Algae at the University of Gottingen, Germany (SAG). Chlorella vulgaris (CCAP 211/19), Scenedesmus communis (CCAP 276/4B) and Tetradesmus obliquus (CCAP 276/10) were obtained from the UK Culture Collection of Algae and Protozoa (CCAP, Scottish Association for Marine Science, Oban, Scotland, U.K.). Stock cultures and further cultivation were maintained on 3 N-BBM +V medium [10]. Each species was cultured in 1L reactors with inline 0.22 µm filters to prevent contamination and aeration of 0.15 vvm. Cultures were grown in triplicate under fluorescent light (100 µmol photons m-2s-1), 16/8 h light/dark for 21 days in batch mode. Biomass was harvested by centrifugation at 3200g, 10 min, RT. Freshly harvested microalgae biomass was frozen at -80°C, solidifying the water inside the cells, then freeze-dried by a vacuum evaporator (Alpha 2-4 LD plus, Martin Christ) at -70°C for 48h, until achieving a final low moisture content (usually <5%).
2.2. Total Lipid Content and Fatty Acid Analysis
Total lipids were extracted using a modified multi-step Bligh & Dyer procedure with chloroform : methanol, followed by phase separation and gravimetric quantification as outlined in [11]. The dried lipid extracts were re-dissolved in n-hexane, with tripentadecanoin (100 μg mL⁻¹) added as an internal standard. Residual moisture was removed using anhydrous sodium sulphate. Transesterification was carried out with 2 M potassium methoxide and 20% HCl in methanol, followed by heating at 70 °C for 30 min and centrifugation at 3200 × g for 10 min at room temperature. The upper n-hexane layer containing fatty acid methyl esters (FAMEs) was filtered through a 0.2 μm PTFE membrane. FAME concentrations were determined by gas chromatography with flame ionisation detection (GC-FID) using a 30 m HP-5 capillary column (Agilent) and helium as the carrier gas (2 mL min⁻¹, 10:1 split ratio). The separation employed a four-step oven program: 50 °C for 2 min, ramping at 10 °C min⁻¹ to 150 °C, then at 17.5 °C min⁻¹ to 185 °C, and finally to 300 °C. The injector and detector temperatures were set at 280 °C and 320 °C, respectively. Fatty acids (FA) were quantified using a 37-FAME standard mixture (Sigma Aldrich, Ireland) and linear calibration curves, with identification based on retention times. Transesterification efficiency was calculated from the conversion rate of the C15:0 internal standard, and this value was applied to correct the fatty acid quantities in each sample [12].
2.3. D. melanogaster Assay
Parental flies of wildtype Canton-S (CS) strain (Bloomington Drosophila stock centre at Indiana University, USA) were kept at 25°C and held on standard fly media (agar 0.25g, yeast 0.375g, sugar 1.25g and semolina 2.0g per 25mL of distilled water) in a climate room with a LD 12 h:12 h cycle and 50% relative humidity. For experimental variants, 3 mg of freeze-dried microalgae biomass was added per 1 mL of fly standard media. The lifespan and body weight were measured separately for females and males. For that, 100 flies of each sex were raised in 250-mL bottles containing 20 mL of medium (20 flies per vial) under the same conditions as noted above. Flies were transferred into fresh food vials with the same component every 4 days. Day 3 post-ecdysis, and the deaths of the flies were recorded. Flies were considered dead when neither voluntary movement nor responses to external stimulation could be observed. The average body weights of flies from the 5 replicated vials were measured using a Denver Instrument SI-234 balance (accuracy 0.0001 g). The experiments stopped on day 45 when most flies died.
To assess fecundity, pairs of D. melanogaster (one male and one female) were placed in vials containing 5 mL of standard nutritional medium for control and medium with added microalgae powder for experimental replicates. All vials were maintained for 72 hours. After this period, the adult flies were removed, and the number of emerging adult offspring (imago) was recorded. The total number of offspring per pair was used as a measure of reproductive output (fecundity).
2.4. Statistical Analysis
The data related to total lipids and FAs concentration presented in this study are reported as mean ±standard deviation
Drosophila survival days estimates were expressed as the median ± percentage difference from the control group as performed in [13]. The body weight of flies and fecundity were expressed as mean ±SEM.
GraphPad Prism 10.4 (Boston, Massachusetts, USA) was used for statistical analysis. A one-way analysis of variance (ANOVA) with post-hoc Tukey's test was applied for comparison of total lipid and total FA content, while Dunnett's test was used for body weight and fecundity. Lifespan comparisons were conducted using the nonparametric Kruskal-Wallis criterion with post hoc Mann-Whitney tests. Differences were considered statistically significant at p-value < 0.05.
3. Results
3.1. Total Lipid and Fatty Acid Profile
The total lipid content and fatty acid (FA) profile of four microalgae species are shown in Table 1.
The total lipid concentration varied among the analyzed microalgae strains, ranging from 16.6 ± 3.0% in S. communis to 27.2 ± 0.9 % in N. limnetica. Lipid content of C. vulgaris and T. obliquus was 26.7 ± 2.8% and 22.6 ± 0.9% respectively. The total fatty acid content was 3.4 and 3.5 times lower than the total lipid content in C. vulgaris and N. limnetica, and 5.7 and 6.2-fold lower in S. communis and T. obliquus. All microalgal species produced unsaturated FAs C15:0, C16:0, as well as monounsaturated fatty acids C18:2n6 (omega-6), and C18:1n9 (omega-9), though in varying proportions. Only trace amounts of additional fatty acids were observed: C14:1 (1.6 ± 0.1%), C18:0 (2.9 ± 0.1%), and C16:1 (3.9 ± 0.2%) in N. limnetica, and C18:1 (4.1 ± 0.2%) and C16:1 (5.4 ± 0.3%) in C. vulgaris. Thus, the most relevant FAs for dietary application identified in microalgae strains were Linoleic acid (C18:2n6C; omega-6) and Oleic acid (C18:1n9; omega-9).
3.2. Effect of Microalgae-Treated Food on D. melanogaster Physiological Parameters
3.2.1. Lifespan
Survival days estimates were expressed separately for females and males, as the percentage of median difference from the control group (Table 2).
In the control group, the median lifespan differed between females and males, at 18 and 12 days, respectively. Inclusion of microalgal freeze-dried biomass at 3 mg/mL extended the female median lifespan by 45.9% for C. vulgaris, 8.1% for S. communis, and 27.0% for T. obliquus. A more significant extension was observed in males, where supplementation with C. vulgaris, N. limnetica, S. communis, and T. obliquus biomass increased their median lifespan by up to 75.0%, 25%, 29.2% and 50.0% respectively.
3.2.2. Body Mass and Fertility
The fly’s body mass, obtained separately for females and males (Figure 1A) and fertility (Figure 2B) are shown below.
The average body mass of D. melanogaster females was 1.23 ± 0.03 mg, and that of males was 0.95 ± 0.09 mg in the control variant. Supplementation with microalgal biomass had no significant effect on their body mass, except in females treated by N. limnetica, where the imago weight decreased by 21% (p < 0.05).
The average progeny from Control D. melanogaster couples was equal to 60.07 ± 2.8. A statistically significant increase of 20% and 18% in fertility was observed only with the addition of T. obliquus and S. communis, respectively, to the nutritional medium.
4. Discussion
Currently, microalgae are regarded as a sustainable source of lipids for diverse applications, ranging from biofuels [3] to nutritional supplements [14,15]. In our study, we investigated the potential application of four oleaginous microalgal strains as a dietary source of essential fatty acids (FAs) using fruit fly D. melanogaster as a model organism for evaluation. In our experiment, after 21 days of autotrophic cultivation in 3N-BBM-V medium with aeration, the highest total lipid concentrations (%DW) were observed for C. vulgaris (26.7 ± 2.8), N. limnetica (27.2 ± 0.9), and T. obliquus (22.6 ± 0.9), with no statistically significant differences among them (Table 1). The lowest lipid concentration was found in S. communis (16.6 ± 3.0). These values are consistent with, or slightly differ from, those reported by other authors. For example, Scenedesmus obliquus (the former name of T. obliquus) showed 19.7 ± 2.1 %DW, and C. vulgaris 27.3 ± 1.2%DW, when cultivated on MBBM medium [9]. Arora et al. [16] reported that C. vulgaris UTEX 395 cultured on BBM achieved a lipid productivity of 33.8 ± 1.2 mg/L/d, which increased to 40% DW when grown on sweet sorghum bagasse hydrolysate under mixotrophic conditions. Nitrogen starvation enhanced lipid content in C. vulgaris ESP-31 up to 55.9% [17] and for C. vulgaris FACHB-1068 up to 31.33% [18], but generally reduced biomass productivity, pigment concentration, and protein content [18] as well as thickened cell walls, lowering cell rupture efficiency [4,19]. The N. limnetica strain, similar to that used in our study, when cultured on OHM media with 10 mM KNO3 under semi-continuous conditions, exhibited a lipid concentration of 24% DW [20]. Fardous et al. [21] mention that the highest lipid concentration for S. communis (20.92% DW) occurred when grown on BBM with the lowest NaNO3 concentration. Overall, microalgal capacity to synthesize lipids, as well as the resulting FA profile, depends strongly on the microalgae strain [16], the composition of cultivation medium [16], and the cultivation conditions [14,16,17]. FA, one of the fractions of total lipids, typically includes free groups. FAs bonded to triacylglycerols (TAGs) serve as energy storage, while those incorporated into phospholipids and glycolipids play structural roles. Only minor amounts exist as free FA molecules [22]. TAGs mainly consist of saturated fatty acids (SFAs) and monounsaturated fatty acids (MUFAs), including C14:0 (myristic acid, MA), C16:0 (palmitic acid, PA), C16:1 (palmitoleic acid, POA), C18:0 (stearic acid, SA), and C18:1 (oleic acid, OA). They may also contain polyunsaturated fatty acids (PUFAs). In particular, long-chain PUFAs are valuable in the nutraceutical and food industries because of their nutritional benefits. Notably, PUFAs such as C20:5 (eicosapentaenoic acid, EPA) and C22:6 (docosahexaenoic acid, DHA) are especially important because there are currently no plant-based sources providing them [3].
FA profile is also highly variable and strongly depends on physical and chemical cultivation parameters [1,23]. Even the same strain growing in variations in light intensity, temperature, nutrient limitation, pH, oxidative stress, osmotic shock, and C:N ratio can cause substantial differences in both the quantity and quality of FAs produced [3,11]. This high degree of variability makes it challenging to directly compare results reported in different studies, but it also highlights promising opportunities for optimizing cultivation conditions to enhance FA production in microalgae.
In our GC assay that contained a 37-FAME standard mixture, neither EPA nor DNA was detected in the observed microalgal strains. The longest-chain unsaturated FA identified were C18:1n9 (omega-9), and C18:2n6 (omega-6), with the highest concentration observed in C. vulgaris and N. limnetica strains. Omega-9 (oleic acid) is a non-essential, associated with anti-inflammatory effects [24]. Omega-6 (linoleic acid) is an essential FA that plays a crucial role in cell structure. Both omega-9 and omega-6 are involved in various bodily functions, but an imbalance in their intake can lead to negative health consequences [25]. Unsaturated FAs C15:0, C16:0 – others that were detected in all the observed microalgae strains. The first one, pentadecanoic acid (C15:0), is an essential odd-chain saturated fatty acid with broad activities relevant to protecting cardiometabolic, immune, and liver health, enhances processes associated with longevity and health span [26]. On the other hand, C16:0 (palmitic acid), an even-chain saturated fatty acid, is often viewed less favourably for health outcomes but can be converted to C15:0 through the α-oxidation process in some bacteria [27]. Thus, the question of whether microalgae can serve as a safe source of lipids for animal and human nutrition remains open. Since D. melanogaster shares over 60% of its genes with humans [6] and possesses several tissues, such as the gut (intestines), oenocytes (liver), and the fat body (adipose tissue) [7], the experiments involving this model organism can help address this research gap.
In current research, 3 mg of freeze-dried microalgal biomass powder was added to 1 mL of standard nutritional medium for feeding D.melanogaster. In this setup, microalgae served as the sole source of lipids. Subsequently, key health parameters, such as lifespan, fertility, and body mass, were evaluated. The most pronounced changes were observed in lifespan, which significantly increased in all experimental groups except in females fed with the addition of N. limnetica biomass. As expected, male flies, whose lifespan is typically shorter than that of females [28], showed a greater improvement. Much fewer changes were observed in the fly body mass and fecundity. Only females fed with N.limnetica biomass showed a slight decrease in body mass (by 21%). Fecundity was enhanced in couples reared on medium supplemented with S.communis and T.obliquus biomass. Overall, dietary supplementation with microalgae at 0.3% of daily food had a positive effect on the health of D. melanogaster. In the literature, information regarding the effect of microalgae supplementation on D. melanogaster physiology is almost absent. To date, only two studies, both by Qiu and colleagues, have been reported. In the first study [9], a relatively high supplementation level (40%) of C. vulgaris and S. obliquus biomass resulted in decreased body weight (0.51 ± 0.006 mg vs. 0.60 ± 0.005 mg for control) and a shorter mean lifespan (36 days vs. 55.8 days for control). In contrast, their subsequent study [7] demonstrated opposite results, where feeding by 4 mg/mL wastewater-cultivated C. sorokiniana significantly extended the median lifespan of Drosophila (55 days vs. 49 days for control). Thus, our study addresses an important gap by evaluating the nutritional value of microalgae using D.melanogaster, a model that remains largely underexplored in this context. The findings provide initial evidence that microalgae supplementation can positively influence key health parameters, underscoring their potential as a sustainable lipid source. Future research should focus on two main directions: (i) enhancing lipid production in the studied microalgal strains through optimization of cultivation conditions, including the use of wastewater as a growth medium, and (ii) extending evaluation in D. melanogaster to a broader range of physiological parameters, with potential implications for advancing human nutritional research.
Author Contributions
Conceptualization, S.M. and R.H.; methodology, S.M., S.B., Y.H.; investigation, S.M., S.B., Y.H.; data curation, S.M.; writing—original draft preparation, S.M.; writing—review and editing, S.M., S.B., R.H.; visualization, S.M., Y.H.; supervision, R.H.; project administration, R.H.; funding acquisition, R.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Irish Research Council, grant number Project: IRC URS/2022/3L “Cell-Wall Autolysis for the Scalable Fractionation of Microalgae into Biofuels and Novel Food Products”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors would like to thank Hans-Georg Eckhardt for technical assistance with GC-FID.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Krienitz, L.; Wirth, M. The High Content of Polyunsaturated Fatty Acids in Nannochloropsis Limnetica (Eustigmatophyceae) and Its Implication for Food Web Interactions, Freshwater Aquaculture and Biotechnology. Limnologica 2006, 36, 204–210. [Google Scholar] [CrossRef]
- Ferreira De Oliveira, A.P.; Bragotto, A.P.A. Microalgae-Based Products: Food and Public Health. Future Foods 2022, 6, 100157. [Google Scholar] [CrossRef]
- Morales, M.; Aflalo, C.; Bernard, O. Microalgal Lipids: A Review of Lipids Potential and Quantification for 95 Phytoplankton Species. Biomass and Bioenergy 2021, 150, 106108. [Google Scholar] [CrossRef]
- Shivakumar, S.; Serlini, N.; Esteves, S.M.; Miros, S.; Halim, R. Cell Walls of Lipid-Rich Microalgae: A Comprehensive Review on Characterisation, Ultrastructure, and Enzymatic Disruption. Fermentation 2024, 10, 608. [Google Scholar] [CrossRef]
- Slocombe, S.P.; Zhang, Q.; Ross, M.; Anderson, A.; Thomas, N.J.; Lapresa, Á.; Rad-Menéndez, C.; Campbell, C.N.; Black, K.D.; Stanley, M.S.; et al. Unlocking Nature’s Treasure-Chest: Screening for Oleaginous Algae. Sci Rep 2015, 5, 9844. [Google Scholar] [CrossRef]
- Staats, S.; Lüersen, K.; Wagner, A.E.; Rimbach, G. Drosophila Melanogaster as a Versatile Model Organism in Food and Nutrition Research. J. Agric. Food Chem. 2018, 66, 3737–3753. [Google Scholar] [CrossRef]
- Musselman, L.P.; Kühnlein, R.P. Drosophila as a Model to Study Obesity and Metabolic Disease. Journal of Experimental Biology 2018, 221, jeb163881. [Google Scholar] [CrossRef]
- Qiu, S.; Shen, Y.; Zhang, L.; Ma, B.; Amadu, A.A.; Ge, S. Antioxidant Assessment of Wastewater-Cultivated Chlorella Sorokiniana in Drosophila Melanogaster. Algal Research 2020, 46, 101795. [Google Scholar] [CrossRef]
- Qiu, S.; Wang, S.; Xiao, C.; Ge, S. Assessment of Microalgae as a New Feeding Additive for Fruit Fly Drosophila Melanogaster. Science of The Total Environment 2019, 667, 455–463. [Google Scholar] [CrossRef]
- Harry W., Bischoff; Harold, C. Bold Phycological Studies IV. Some Soil Algae from Enchanted Rock and Related Algal Species; Department of Biology, Texas Lutheran College, Seguin, Texas.; 1963; Vol. No. 6318.
- Halim, R.; Papachristou, I.; Kubisch, C.; Nazarova, N.; Wüstner, R.; Steinbach, D.; Chen, G.Q.; Deng, H.; Frey, W.; Posten, C.; et al. Hypotonic Osmotic Shock Treatment to Enhance Lipid and Protein Recoveries from Concentrated Saltwater Nannochloropsis Slurries. Fuel 2021, 287, 119442. [Google Scholar] [CrossRef]
- Kiani, H.; Ma, Q.; Xiao, M.; Li, Y.; Brooke, F.J.; Mulcahy, S.; Miros, S.; Halim, R. Growth and Fatty Acid Profile of Nannochloropsis Oceanica Cultivated on Nano-Filtered Whey Permeate. J Appl Phycol 2024, 36, 2503–2516. [Google Scholar] [CrossRef]
- Liubertas, T.; Poderys, J.; Vilma, Z.; Capkauskiene, S.; Viskelis, P. Impact of Dietary Potassium Nitrate on the Life Span of Drosophila Melanogaster. Processes 2021, 9, 1270. [Google Scholar] [CrossRef]
- Freire, I.; Cortina-Burgueño, A.; Grille, P.; Arizcun Arizcun, M.; Abellán, E.; Segura, M.; Witt Sousa, F.; Otero, A. Nannochloropsis Limnetica: A Freshwater Microalga for Marine Aquaculture. Aquaculture 2016, 459, 124–130. [Google Scholar] [CrossRef]
- Tiwari, B.K.; Garcia-Vaquero, M. Nutraceuticals from Algae: Current View and Prospects from a Research Perspective. Marine Drugs 2022, 20, 671. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Philippidis, G.P. Insights into the Physiology of Chlorella Vulgaris Cultivated in Sweet Sorghum Bagasse Hydrolysate for Sustainable Algal Biomass and Lipid Production. Sci Rep 2021, 11, 6779. [Google Scholar] [CrossRef] [PubMed]
- Yeh, K.; Chang, J. Nitrogen Starvation Strategies and Photobioreactor Design for Enhancing Lipid Content and Lipid Production of a Newly Isolated Microalga Chlorella Vulgaris ESP-31: Implications for Biofuels. Biotechnology Journal 2011, 6, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chen, Z.; Xiao, Y.; Yuan, M.; Zhou, C.; Liu, G.; Fang, J.; Yang, B. Biochemical and Morphological Changes Triggered by Nitrogen Stress in the Oleaginous Microalga Chlorella Vulgaris. Microorganisms 2022, 10, 566. [Google Scholar] [CrossRef]
- Rahimi, M.; Webley, P.A.; Martin, G.J.O.; Halim, R. Nitrogen Starvation for Fuel Production from Nannochloropsis: A Trade-off between Calorific Lipid Accumulation and Energy Loss for Cell Disruption. Chemical Engineering Journal Advances 2025, 23, 100812. [Google Scholar] [CrossRef]
- Freire, I.; Cortina-Burgueño, A.; Grille, P.; Arizcun Arizcun, M.; Abellán, E.; Segura, M.; Witt Sousa, F.; Otero, A. Nannochloropsis Limnetica: A Freshwater Microalga for Marine Aquaculture. Aquaculture 2016, 459, 124–130. [Google Scholar] [CrossRef]
- Fardous Ara Mukta; Helena Khatoon; Mohammad Redwanur Rahman; Mahima RanjanAcharjee; Subeda Newase; Zannatul Nayma; Razia Sultana; Shanur Jahedul Hasan Effect of Different Nitrogen Concentrationson the Growth, Proximate and Biochemical Composition of Freshwater Microalgae Scenedesmus Communis. Journal of Energy and Environmental Sustainability 11 & 12, 36–42.
- Shen, P.-L.; Wang, H.-T.; Pan, Y.-F.; Meng, Y.-Y.; Wu, P.-C.; Xue, S. Identification of Characteristic Fatty Acids to Quantify Triacylglycerols in Microalgae. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Breuer, G.; Lamers, P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. Effect of Light Intensity, pH, and Temperature on Triacylglycerol (TAG) Accumulation Induced by Nitrogen Starvation in Scenedesmus Obliquus. Bioresource Technology 2013, 143, 1–9. [Google Scholar] [CrossRef]
- Farag, M.A.; Gad, M.Z. Omega-9 Fatty Acids: Potential Roles in Inflammation and Cancer Management. Journal of Genetic Engineering and Biotechnology 2022, 20, 48. [Google Scholar] [CrossRef]
- Johnson, M. Omega-3, Omega-6 and Omega-9 Fatty Acids: Implications for Cardiovascular and Other Diseases. J Glycomics Lipidomics 2014, 04. [Google Scholar] [CrossRef]
- Venn-Watson, S.; Schork, N.J. Pentadecanoic Acid (C15:0), an Essential Fatty Acid, Shares Clinically Relevant Cell-Based Activities with Leading Longevity-Enhancing Compounds. Nutrients 2023, 15, 4607. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, B. The Relative Contribution of Propionate, and Long-Chain Even-Numbered Fatty Acids to the Production of Long-Chain Odd-Numbered Fatty Acids in Rumen Bacteria. Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism 1978, 528, 239–246. [Google Scholar] [CrossRef]
- Lints, F.A.; Bourgois, M.; Delalieux, A.; Stoll, J.; Lints, C.V. Does the Female Life Span Exceed That of the Male. Gerontology 1983, 29, 336–352. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
D. melanogaster body mass (A) and fertility (B) measurement after feeding with microalgae freeze-dried biomass (3 mg/mL). Ch (C. vulgaris), NL (N. limnetica), Sc (S. communis), TO (T. obliquus). Comparisons for parameters between microalgae-treated flies against control flies were performed by one-way ANOVA with post-hoc Dunnett's test. Values are means ±SD. **** – p-value < 0.0001, *** - p-value < 0.01, ** – p-value < 0.05 (n = 5).
Figure 1.
D. melanogaster body mass (A) and fertility (B) measurement after feeding with microalgae freeze-dried biomass (3 mg/mL). Ch (C. vulgaris), NL (N. limnetica), Sc (S. communis), TO (T. obliquus). Comparisons for parameters between microalgae-treated flies against control flies were performed by one-way ANOVA with post-hoc Dunnett's test. Values are means ±SD. **** – p-value < 0.0001, *** - p-value < 0.01, ** – p-value < 0.05 (n = 5).
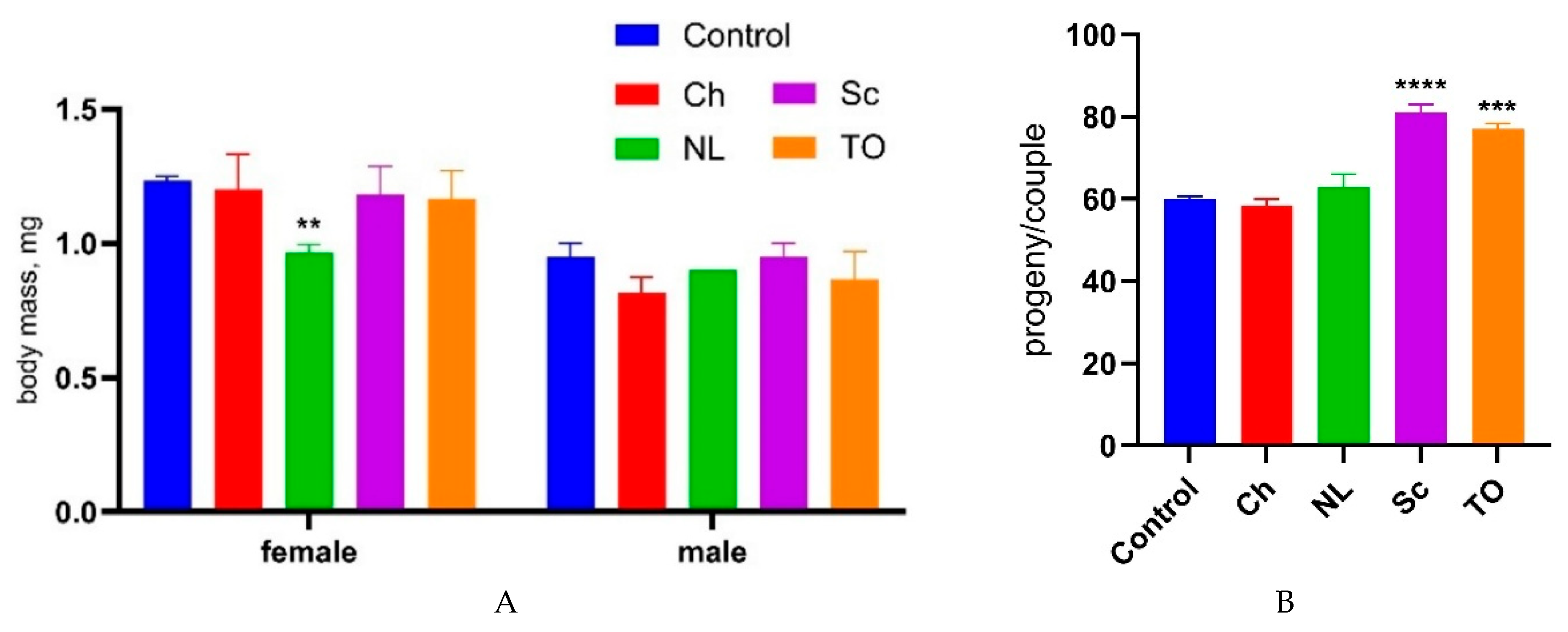

Table 1.
The total lipid content and fatty acid profile of microalgae grown on standard medium.
| Component, % DW | C. vulgaris | N. limnetica | S. communis | T. obliquus |
|---|---|---|---|---|
| Total Lipids | 26.7 ± 2.8A | 27.2 ± 0.9A | 16.6 ± 3.0B | 22.6 ± 0.9AB |
| Total Fatty Acids | 8.0 ± 0.9A | 7.7 ± 0.7AB | 2.9 ± 0.8BC | 3.7 ± 1.6C |
| Fatty acids, (% TFA) | ||||
| Saturated | ||||
| C14:0 | – | 1.6 ± 0.1 | – | – |
| C15:0 | 16.2 ± 7.6 | 15.9 ± 5.8 | 41.7 ±13.0 | 38.4 ± 12.1 |
| C16:0 | 17 ± 1.5 | 14.9 ±1.4 | 13.8 ± 6.3 | 11.9 ± 2.1 |
| C18:0 | 4.1 ± 0.2 | 2.9 ± 0.1 | – | – |
| Monounsaturated | ||||
| C16:1 | 5.4 ± 0.3 | 3.9 ± 0.2 | – | – |
| C18:1n9 Omega-9 | 46.0 ± 0.6 | 43.7 ± 1.2 | 28.9 ± 4.1 | 41.0 ± 0.6 |
| C18:2n6 Omega-6 | 15.9 ± 0.4 | 21.2 ± 0.2 | 15.5 ± 2.7 | 17.3 ± 2.8 |
A, B letters indicate statistical grouping based on a one-way ANOVA with post-hoc multiple comparisons using Tukey's test (p<0.05). Groups that do not share the same letter are significantly different. Values are means ± SD (n=2).
Table 2.
Lifespan of D. melanogaster following dietary supplementation with freeze-dried microalgal biomass.
Table 2.
Lifespan of D. melanogaster following dietary supplementation with freeze-dried microalgal biomass.
| Variant | Females | Males | ||||||
|---|---|---|---|---|---|---|---|---|
| Survival Days | Median lifespan change (% vs. control) |
Survival Days | Median lifespan change (% vs. control) |
|||||
| Median | Mean | Max | Median | Mean | Max | |||
| Control | 18.5 | 17,2 | 35 | 0A | 12 | 13.8 | 30 | 0A |
| C. vulgaris | 27.0 | 24.5 | 41 | +45.9B | 21 | 18.9 | 32 | +75.0B |
| N. limnetica | 18.5 | 18.9 | 32 | 0.0A | 15 | 15.3 | 30 | +25.0B |
| S. communis | 20.0 | 20.0 | 36 | +8.1B | 15.5 | 15.0 | 29 | +29.2B |
| T. obliquus | 23.5 | 24.1 | 44 | +27.0B | 18 | 16.8 | 31 | +50.0B |
A, B letters indicate statistical grouping based on nonparametric Kruskal–Wallis criterion with a post-hoc Mann–Whitney test (p<0.05) within females and males separately. Groups that do not share the same letter as the control are significantly different.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



