
Submitted:
14 August 2025
Posted:
14 August 2025
You are already at the latest version
Abstract
Seed and pod decay significantly threaten to soybean yield and quality during the fruiting stage worldwide. Although, few previous studies partially explained the occurrence of Fusarium species on soybean seeds and pods, However, fungal diversity affecting soybean pods in Sichuan Province China, third-largest soybean cultivation region remains unknown. In this study, we identified pod-infecting fungal communities and evaluated their pathogenic potential on soybean seeds and pods. Using morphological indices and DNA barcode markers, we characterized Fusarium species, including F. verticillioides, F. incarnatum, F. equiseti, F. proliferatum, F. fujikuroi, F. oxysporum, F. chlamydosporum, and F. acutatum through Translation elongation factor gene (EF1-α) and RNA polymerases II second largest subunit (RPB2) gene analysis. Multi-locus phylogeny assay of Internal transcribed spacer (rDNA ITS), β-tubulin (β-tubulin), Glyceraldehyde 3-phosphate dehydrogenase (GADPH), Chitin Synthase 1 (CHS-1), Actin (ACT), Beta-tubulin II (TUB2) and Calmodulin (CAL) distinguished Colletotrichum species as C. truncatum, C. karstii, C. cliviicola, C. plurivorum, C. boninense, and C. fructicola. Pathogenicity assays revealed significant damage from Fusarium and Colletotrichum isolates on soybean pods and seeds, with varying isolation frequencies. Among them, F. proliferatum, F. acutatum, and F. verticillioides caused the most severe symptoms, while C. fructicola was most pathogenic, followed by C. truncatum, C. karstii, C. cliviicola, C. plurivorum, and C. boninense. These findings highlight emerging virulent pathogens responsible for soybean pod decay, paves a valuable foundation for developing resistant cultivars to manage pod associated diseases at the later growth stage of soybean.
Keywords:
soybean pods mycoflora
; fungal diversity
; morpho-molecular phylogeny
; pathogenicity assay
; disease management
1. Introduction
Soybean (Glycine max L.), is known as a globally paramount legume crop cultivated for millennia [1,2,3]. It serves as a critical source of plant-based protein (40%) and oil (20%) for human nutrition and animal feed, underpinning global food security [4,5]. Nevertheless, seed-borne diseases caused by multiple hazardous pathogens during the reproductive stages inflict severe economic losses through substantial yield reduction, poor seed quality, and compromised marketability [6]. These pathogens, such as Fusarium, Colletotrichum, Diaporthe, Sclerotinia, Cercospora, Phytophthora, have been predominantly reported to infect various soybean organs [7,8]. Among the most detrimental pathogens, Fusarium and Colletotrichum stand out due to their widespread prevalence and destructive impact on seed development and viability [9,10,11,12].
Fusarium sp. are ubiquitous seed-borne pathogens responsible for complex diseases including root rot, pod blight, seed rot, and sudden death syndrome, that severely diminish germination rates and seedling vigor worldwide [13,14,15,16]. Their genetic diversity and adaptability have been well documented, with distinct species exhibiting pronounced pathogenicity across different regions. For instance, Fusarium proliferatum displays high aggressiveness in China’s Hubei province [12], whereas F. oxysporum, F. equiseti, and F. graminearum are particularly pathogenic in Sichuan province [7,17,18,19,20,21]. Critically, these fungi frequently exist as single entities but as pathogen complexes, greatly complicating disease management strategies and underscoring the urgent need for precise identification at the species level [22,23,24]. A wide spectrum of Fusarium species, such as F. solani, F. oxysporum, F. acuminatum, F. avenaceum, F. cerealis, F. culmorum, F. equiseti, F. graminearum, F. proliferatum, F. pseudograminearum, F. fujikuroi, F. asiaticum, F. commune, and F. verticillioides, has been associated with soybean isolated from various tissues [25,26,27,28,29,30,31,32]. Numerous studies investigating cultivar resistance, pathogenicity, distribution and incidence rates provide evidences that the colonization of soybean roots by multiple Fusarium species is commonplace. However, significant knowledge gaps persist regarding their precise roles and pathogenicity dynamics specifically affecting soybean seeds and pods, as well as the ultimate impacts on seed quality and yield [30,33]. Similarly, Colletotrichum sp. ranking among the top 10 most significant plant pathogenic fungi globally, pose a substantial threat. Their exceptionally broad host range (infecting over 3 000 plant species) and capacity for latent infections make effective control exceptionally challenging [34,35,36]. In soybean, Colletotrichum sp. can induce anthracnose across all developmental stages [37], and symptoms manifest as leaf spotting, stem lesions, pod necrosis, and premature defoliation, collectively contributing to considerable yield losses [38]. Crucially, this disease is primarily seed-transmitted, and infected seeds lead to damping-off of seedlings and the development of lesions on cotyledons during the V1 and V2 developmental stages [39]. The genetic diversity within this pathogen complex, along with key epidemiological and biological characteristics of its constituent members, remains poorly characterized, thus necessitating more precise and comprehensive studies [40,41,42,43,44]. Importantly, for both Fusarium and Colletotrichum pathogens, accurate identification at the species level is the indispensable foundation for developing effective disease management.
Traditional morphological methods often prove inadequate due to overlapping characteristics within species complexes [45,46]. Consequently, molecular approaches, particularly multi-locus phylogenetic analysis targeting conserved genes, such as Internal transcribed spacer (rDNA ITS), Translation elongation factor gene (EF1-α), RNA polymerases II second largest subunit (RPB2) for Fusarium isolates, and β-tubulin (β-tubulin), Glyceraldehyde 3-phosphate dehydrogenase (GADPH), Chitin Synthase 1 (CHS-1), Actin (ACT), Beta-tubulin II (TUB2) and Calmodulin (CAL) for Colletotrichum isolates [47,48,49], have become the important standard for robust species delineation and understanding population dynamic; thereby, such precision is vital for tracking the emerging isolates and designing targeted interventions.
Intercropping systems are widely adopted due to their efficient utilization of light resources, improvement of soil structure through microbes, significant reduction of weeds and pests, and higher production under eco-friendly conditions compared to monoculture systems [21]. In Southwest China, particularly Sichuan province, the widespread adoption of maize-soybean relay strip intercropping enhances land productivity (high Land Equivalent Ratio, LER>1.5), soil health, resource use efficiency, disease and pest suppression [50,51]. However, the characteristic high humidity, moderate temperatures, and limited sunlight in this region create a microenvironment highly conducive to fungal proliferation, infection and dispersal [52,53]. Studies confirm that such cool, humid highland conditions significantly shape pathogenic fungal communities and intensify disease pressure, posing a persistent threat to soybean production [54]. Emerging evidence suggests pathogen populations exhibit rapid adaptation, complex genetic diversity, and increased aggressiveness, potentially linked to changing climatic conditions and agricultural practices [55]. Despite this, comprehensive data on the diversity and pathogenicity of seed- and pod- associated fungi, particularly within the distinctive intercropping pattern in Sichuan province, remain limited. Therefore, this study aims to systemically isolate and characterize the mycobiota associated with soybean seeds focusing on Fusarium and Colletotrichum species based on multi-gene phylogenetic analysis, and additionally, the in vitro pathogenicity of typically dominant Fusarium and Colletotrichum isolates. The findings of the current study offered a promising way to elucidate composition and threat level of major seed pathogens in a critical soybean-growing region with distinctive intercropping cultivation and are expected to provide a useful scientific foundation for typical disease management and soybean resistance breeding.
2. Materials and Methods
2.1. Sampling and Fungal Isolation
A survey was conducted at experimental site to collect soybean pods with discoloration, decay, and the presence of mycelium, under maize-soybean relay strip intercropping from five different soybean cultivating regions in Sichuan province, Southwest China. The fungal pathogens were isolated by washing pod samples under running tap water, followed by drying, excising into small fragments (4-8 mm), and surface sterilization using 1% sodium hypochlorite (w/v) for 1 minute and 75% ethanol (w/v) for 2 minutes. The small fragments were washed thrice, dried on sterile filter paper, and transferred to potato dextrose agar plates (PDA; potato 200 g·L-1, glucose anhydrous 10 g·L-1, and agar 15 g·L-1)[25]. The plates were incubated for 7-15 days at 25±2°C in complete darkness, and the fungal isolates were purified by transferring active marginal hyphae onto fresh PDA plates.
2.2. Morphological Identification of Fungal Population
The morphological identification of fungal population was executed by observing colony feature and texture, the length and width of macroconidia, number of septa, conidial shape and size, and growth rate. These fungal isolates were incubated at 25±2°C and colony growth was recorded after two days of incubation, while the growth rate of Fusarium species were documented after 7 days of incubation in dark conditions. For Colletotrichum species morphological identification, except growth rate (two days of post inoculation) the other cultural variables were distinguished after 15 days of post-incubation. In addition, the general PDA and species-specific CMC (Carboxymethyl-Cellulose 15.0 g·L-1, KH2PO4 monobasic 1.0 g·L-1, NH4NO3 1.0 g·L-1, yeast extract 1.0 g·L-1, MgSO4.7H2O 0.5 g·L-1 in distilled water) media were used for the conidial spore production of Fusarium species. Similarly, PDA medium was utilized to generate enough spores of Colletotrichum species accordingly. The number and size of conidia (n=50) were recorded carefully of each fungal species by observing them under the accessible compound microscope (Eclipse 80i, Nikon, Japan) [56].
2.3. Molecular Identification of Fungal Genera
The 7-day-old mycelium of fungal isolates cultured on PDA plates were scraped with disinfected blades to extract the genomic DNA, following the standard manual of the Rapid Fungi Genomic DNA Isolation Kit (Sangon Biotech, Shanghai, China). The quality and quantity of DNA were assessed using a NanoDrop™ 2000 Spectrophotometer (Thermo Scientific, Delaware, USA) after extraction. For each fungal genus, specific primer pairs (listed in Table S1) were selected for PCR amplification. A 50 µL of reaction mixture was prepared, containing 2 μL of each primer, 25 μL of Taq PCR Master mix, 2 μL of DNA from each isolate, and 19 μL of sterilized water. The Peltier Thermal Cycler (S-1000TM, Bio-Bri, China) was used for amplification, and the temperature conditions for different primers are mentioned in Table S1. The amplified products were subjected to electrophoresis on a 1.0% (w/v) agarose gel in 1×TAE buffer, and samples were sequenced using the ABI-PRISM 3730 automatic sequencer (Applied Biosystems, Foster, USA).
2.4. Phylogenetic Analysis
To evaluate the genetic differences among different fungal genera, molecular evolutionary and phylogenetic analysis were executed. The amplified sequences were trimmed with BioEdit software (developed by Tom Hall; BioEdit free download v.7.0.5.3) and then blasted against multiple databases, including FUSARIUM-ID and Fusarium MLST for Fusarium species [57], and the National Centre for Biotechnology Information (NCBI) for Colletotrichum species. Additionally, Clustal X 1.83 was used for alignment of sequences of each constructed tree by removing gaps (missing barcode information) and weighing the characters universally. Phylogenetic trees of combined barcode for Fusarium and Colletotrichum species were constructed accordingly using MEGA version X with the Neighbor-Joining method, supported by the Tamura–Nei model, respectively [58]. The constructed tree clades were supported by 1000 bootstrap replicates, and resulted sequences were deposited in the NCBI GenBank and TreeBASE (www.treebase.org).
2.5. Pathogenicity Test of Isolated Fungi
For pathogenicity tests, seeds of the soybean cultivar ‘Nandou12’ and soybean pods were inoculated with spore suspension respectively of each fungal species to fulfil Koch's postulates, following the method described by [59] with minor modifications. Three-prototype isolates of each species were selected randomly to analyses their pathogenic impact on seeds and pods respectively. Fusarium spores were produced by adding 3-5 mycelial discs to 20 mL of PDA or CMC medium and then incubated in orbital shaker at 150 r·min−1 at 25°C for 7 days. Similarly, Colletotrichum isolates spores were obtained by scraping fungal mycelium in deionized water. The final spore concentration was adjusted to 1 × 105 spore mL−1 with double distilled water (ddH2O) for inoculation. The seeds and pods were subjected to surface sterilization with 1% NaClO and rinsed three times for 1 minute, followed by air drying under sanitized conditions on double-layered filter paper [60]. Three separate replicates were prepared, each containing 15 seeds per plate and 3 pods for each isolate. The seeds and pods were dipped in spore suspension for 15 minutes to allow diseases development [45]. Seeds treated with ddH2O served as the negative control, and all plates were incubated in the dark at 25 ± 2°C for 7 days with 70% relative humidity. After incubation, disease severity index (DSI) was assessed as described by [8] with minor modifications. In addition, the percentage of mycelium coverage area (PMC) and seed and pod weight were noted. The DSI and PMC were calculated using the following formula:

2.6. Isolation Frequency Analysis of Fungal Isolates
The isolation frequency of fungal genera especially Fusarium and Colletotrichum isolates were calculated by dividing the number of times a specific fungus is isolated by the total number of fungal isolates obtained in the study, then multiplying by 100 to express the result as a percentage. The isolation frequency was deliberated using the following formula:
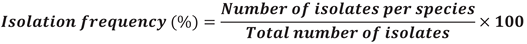
2.7. Data Process and Analysis
The recorded data was processed through Microsoft office excel 2016 (Microsoft Corporation, Redmond, Washington, USA). The DSI and PMC average values were calculated from independent triplicates in pathogenicity tests. In addition, seed and pod weight were subsequentially recorded for each representative isolates of Fusarium and Colletotrichum species. The statistical analysis was performed by applying the Duncan’s multiple range test in IBM SPSS Statistics 20 (IBM Corp., Armonk, N.Y., USA) to underpin the significant differences (P = 0.05).
3. Results
3.1. Identification of Fungal Species Associated with Intercropped Soybean Pods
In the present study, soybean pods (n=182) were collected from soybean under maize-soybean strip intercropping pattern, and total 132 isolates were obtained. Upon morphological characteristics including colonial color and texture of colonies and mycelium, these isolates were primarily clustered into ten groups (Figure 1A). Furthermore, rDNA ITS fragments were amplified and sequenced, thus BLASTn analysis revealed these isolate groups represented ten distinct genera, including Fusarium sp., Colletotrichum sp., Phomopsis/Diaporthe sp., Bipolaris sp., Nigrospora sp., Graphium sp., Clonostachys sp., Nadulisporium sp., Alternaria sp., and Boeremia sp. Phylogenetic analysis showed that different fungal genera were clearly separated but clustered with their corresponding reference isolates (Figure 1B, Table S2).
To analyze the isolation frequency of these fungal genera associated with soybean pods, we found that Fusarium sp. (32.57%) was frequently isolated from intercropped soybean pods, followed by Colletotrichum sp. (28.03%), Phomopsis/Diaporthe sp. (12.12%), Bipolaris sp. (12.12%). Nigrospora sp. (4.54%), Graphium sp. (3.03%), Clonostachys sp. (3.03%), and Nadulisporium sp. (1.51%). Both Alternaria sp. and Boeremia sp. had the lowest isolation frequency accounting for 0.75% (Figure 1C). Thus, fungal communities associated with soybean pods in Sichuan province are well-characterized, and Fusarium and Colletotrichum were the dominant genera among them.
3.2. Identification of Fusarium Species Associated with Soybean Pods
To further verify Fusarium species, morphological features of colonies and conidial spores of different Fusarium isolates were observed as shown in Figure 2A and Table 1. After seven days of incubation, Fusarium isolates displayed a pronounced variation in colony colors, which ranged from filthy white, dark violet and light violet to light pink. The mycelia also exhibited sparse and fluffy white mycelia, dense white or white-purple mixtures. Almost all Fusarium isolates produced pointed and sickle-shaped macroconidia. Based on conidial morphology, size and colony pigmentation, total 43 Fusarium isolates clustered into eight morphological groups.
For molecular validation, the EF1-α and RPB2 genes were amplified, sequenced, and blasted against the Fusarium MLST and FUSARIUM-ID databases. Sequence similarity analysis identified eight distinct Fusarium species: F. verticillioides, F. incarnatum, F. equiseti, F. proliferatum, F. fujikuroi, F. oxysporum, F. chlamydosporum, and F. acutatum. For phylogenetic analysis, maximum-likelihood tree was constructed using combined EF1-α and RPB2 genes. The trees included 43 Fusarium isolates from this study and 17 reference isolates and Nactriaceae sp. (JF740999.1) serving as an outgroup (Table S3). Phylogenetic analysis clearly resolved the taxonomic relationships and genetic distances among the Fusarium species (Figure 2B). All eight species formed distinct clades, except within two species complexes: the F. incarnatum-equiseti complex (FEIC) and the F. proliferatum-fujikuroi complex (FPFC). These species complexes grouped in the same major clade but formed well-supported distinct subclades. Bootstrap support values exceeded 92% for all species and species complex branches. The generated sequences were deposited in GenBank, and accession numbers are provided in Table S4.
3.3. Identification of Colletotrichum Species Associated with Soybean Pods
Morphological analysis of 132 fungal isolates identified 37 isolates as Colletotrichum species. Colonies exhibited white, cottony mycelia, while conidia varied in shape (fusiform, cylindrical and oval/ellipsoidal) presented in Figure 3A and Table 2. There isolates were classified into six morphological groups. Furthermore, Colletotrichum species was confirmed using a six-locus molecular approach including rDNA ITS, CHS, GAPDH, ACT, CAL and TUB2 genes. BLASTn analysis revealed maximum sequence similarity with six distinct Colletotrichum species. Phylogenetic analysis based on these loci employed maximum parsimony and ML methods (1000 bootstrap replicates). The tree included 37 isolates from this study, 12 reference isolates and Monilochaetes infuscans (CBS:869.96) as an outgroup (Table S5). Our results demonstrated that all isolates clustered within a single major clade but resolved into six well-supported species: C. truncatum, C. karstii, C. cliviicola, C. plurivorum, C. boninense, and C. fructicola and their accession numbers were obtained from GenBank (Figure 3B, Table S6).
3.4. Isolation Frequency of Fusarium and Colletotrichum Species
For isolation frequency, F. proliferatum (20.93%), F. fujikuroi (16.27%) and F. equiseti (16.27%) were most prevalent, followed by F. acutatum (13.95%), F. verticillioides (9.3%) and F. incarnatum (9.3%) among Fusarium genera. Compared to other species, F. oxysporum and F. chlamydosporum were the least frequent, with the isolation frequency of 6.97% each (Figure 4A). Among Colletotrichum species, C. fructicola was predominant (21.62%), followed by C. truncatum and C. karstii (18.91% each), C. cliviicola and C. plurivorum (16.21% each). Colletotrichum boninense was the least isolated and accounts for 10.81% of total Colletotrichum isolates (Figure 4B). Hence, the F. proliferatum and C. fructicola were dominant species isolated from soybean pods in Southwest China.
3.5. Pathogenicity of Fusarium Species on Soybean Pods and Seeds
Pathogenicity assays were conducted to evaluate the effects of Fusarium species on soybean pods (Figure 5 and Table 3) and seeds (Figure S1 and Table S7). As shown in Figure 5, all Fusarium species successfully penetrated soybean pods, causing varying degrees of internal seed decay. Among them, F. acutatum and F. verticillioides (100%) resulted in complete PMC (100%), followed by F. proliferatum (90.66%), F. equiseti (88.33%), F. oxysporum (55%), F. fujikuroi (46.66%), F. incarnatum (26.66%), and F. chlamydosporum (23.33%). Interestingly, F. proliferatum, F. acutatum, and F. verticillioides exhibited a DSI of 100%, while F. oxysporum had a DSI of 83.33%, F. fujikuroi, F. chlamydosporum, and F. equiseti all had a DSI of 75%. F. incarnatum showed the lowest DSI (33.33%). Additionally, infected pods exhibited a reduced weight compared to un-inoculated controls, likely due to mycelial overgrowth (Table 1). Internal seed rot was observed with F. proliferatum and F. fujikuroi, while other species caused external rot with minimal discoloration compared to control pods. Seven days post-inoculation, the seeds showed partial to complete coverage by white mycelium (with noted color variation), correlating with species-specific virulence (Figure S1, Table S7). Our results demonstrate that F. proliferatum, F. acutatum and F. verticillioides were the most virulent species.
3.6. Pathogenicity of Colletotrichum Species on Soybean Pods and Seeds
All Colletotrichum species caused diseases on soybean pods, characterized by rotted pods, discolored, and abundant mycelial coverage on pods (Figure 6). All representative isolates of Colletotrichum species resulted in 100% disease incidence on soybean pods (Table 4). Colletotrichum karstii (91.66%) C. fructicola (90%) exhibited the highest PMC, followed by C. truncatum and C. cliviicola (both 81.66%). In contrast, C. boninense (11.66%) and C. plurivorum (10%) showed minimal PMC. DSI was highest for C. fructicola (100%), followed by C. truncatum and C. karstii (both 83.33%), C. cliviicola (66.66%), and C. plurivorum and C. boninense (both 33.33%). Similarly, pod weight varied significantly upon species. C. boninense recorded the highest weight (2.98 g), followed by C. karstii (2.67 g), C. plurivorum (2.59 g), C. cliviicola (2.21 g), C. truncatum (2.16 g), and C. fructicola (2.09 g). Furthermore, internal seed rot with discoloration occurred in pods infected by C. fructicola, C. truncatum, C. karstii and C. cliviicola, while C. plurivorum and C. boninense caused only external discoloration externally. Based on disease severity metrics and symptoms, C. fructicola emerged as the most virulent pathogen on soybean pods (Figure 6). Similarly, Colletotrichum species also caused severe damage to inoculated soybean seed (Figure S2), with all species covering seeds with mycelium. Among these species, representative isolates of C. fructicola had the highest PMC (100%), followed by C. cliviicola (96.66%) and C. boninense (93.33%). However, maximum DSI occurred in C. fructicola and C. truncatum (both 91.66%), trailed by C. boninense (81.66%) and C. cliviicola (70%). Comparably, C. karstii and C. plurivorum exhibited the lowest pathogenicity, with DSI of 58.33% and 38.33%, respectively. In contrast, C. plurivorum and C. boninense caused only external damage with slight discoloration (Table S8). Thus, C. fructicola was the most virulent species on both soybean pods and seeds.
4. Discussion
It is well known that fungal diseases causing soybean seed and pod deterioration significantly reduce global yield and quality [61]. The current study employed morpho-molecular characterization and multi-locus phylogenetic analysis to investigate fungal pathogens in Sichuan province, Southwest China. And we focused on two major genera, Colletotrichum and Fusarium species. Several previous studies documented the occurrence and pathogenic effects of individual Fusarium species [8,11,25,62], our work expands understanding by characterizing the fungal diversity across both genera. We identified eight Fusarium species (F. verticillioides, F. incarnatum, F. equiseti, F. proliferatum, F. fujikuroi, F. oxysporum, F. chlamydosporum, and F. acutatum) and confirmed the presence of the FPFC and FEIC species complexes (Figure 2), aligning with reports from Sichuan province [8]. Additionally, Colletotrichum, the 8th most devastating and wide-spectrum plant pathogen [35], causes anthracnose in soybean and related legumes [63]. We identified six species including C. truncatum, C. karstii, C. cliviicola, C. plurivorum, C. boninense, and C. fructicola (Figure 3).
Furthermore, accurate pathogen identification is essential for disease management [40]. Advances in fungal disease identification have significantly improved the ability to identify a range of pathogenic plant fungi employing specific gene sequence analysis and improved molecular techniques [64,65]. It is well-established that both Fusarium and Colletotrichum species often appear in complexes, sharing similar morphological characteristics (e.g., colony and spore shape) worldwide [66]. To characterize these species, the amplification of two or more genes has emerged as a standard method for accurately identifying specific fungal species within widely spread genera [67]. By witnessing morphological features, molecular procedures, and phylogenetic analysis of RPB2 and EF1-α gene sequences, we recognized 43 different Fusarium species. It has been assumed that two sequencing sections, EF1-α and RNA polymerase largest subunit RPB1 and/or RPB2, are indispensable for Fusarium species characterization [68]. multiple recent studies have used these regions for the precise identification of Fusarium isolates complexes [9,23,24,69]. For Colletotrichum species identification, we used multilocus analysis of ACT, CHS, ITS, GAPDH, TUB2 and CAL genes. employing these genes, we recognized six different Colletotrichum species, including C. boninense, C. truncatum, C. cliviicola, C. karstii, C. fructicola and C. plurivorum. A parallel method has been used to identify the Colletotrichum isolated infecting olive trees and Tea-Oil Camellia (Camellia oleifera C. Abel) [25,26].
Pathogenicity evaluation exposed tissue-specific virulence. Among Fusarium isolates, F. acutatum and F. verticillioides exhibited the highest aggressiveness (PMC and DSI) on soybean seeds. Earlier studies have revealed that F. verticillioides has the ability to diminish soybean seed quality [52], while F. acutatum causes root rot and crown , leading to yellowing and plant death in tomato plants [27]. Among Colletotrichum isolates, C. truncatum and C. fructicola were the furthermost virulent isolates when inoculated on soybean seeds. Interestingly, few past studies have pronounced the C. fructicola as a non-host specific pathogen proficient of infecting an extensive range of plants and crops, including blueberry [22], sugarcane [29], and apple [70]. Except for C. truncatum and C. fructicola, our results presented that C. karstii acted as a moderately aggressive pathogen toward soybean pods and seeds. In line with our study, C. karstii has been reported to cause anthracnose in soybean in China [71]. Many studies have documented that soybean pod diseases lead to seed deterioration, which negatively affects seed germination and reduces grain yield in soybean fields [72]. The pathogenicity outcomes from current study confirmed that the isolated Fusarium and Colletotrichum isolates are lethal and capable of decaying soybean pods and seeds, directly impacting PMC, DSI and seed weight. We predict that F. acutatum, F. verticillioides, C. fructicola, and C. truncatum are destructive plant pathogens responsible for soybean pod and seed decay. Therefore, understanding their infection mechanisms through molecular analysis provides critical insights for developing targeted management strategies.
5. Conclusions
This study underpins the significance of managing fungal pathogens in soybean pods to protect yields. The infected pods from five different locations in Sichuan Province, Southwest China, yielded Colletotrichum and Fusarium species. Through morphological indices and molecular markers analysis, we identified 43 Fusarium isolates. Additionally, six other molecular markers were amplified to identify 37 Colletotrichum isolates. Pathogenicity tests highlights that F. verticillioides, F. acutatum and F. proliferatum were the most destructive, causing noteworthy pod and seed damage, and reduced the seed vigor. Among Colletotrichum isolates, C. fructicola and C. truncatum were the highly aggressive by exhibiting severe symptoms on soybean seeds and pods. These findings underscore the necessity of developing resistant soybean varieties through targeted breeding, particularly against the most aggressive fungal species in high-risk regions like Southwest China, where environmental conditions favor pathogen proliferation, and would greatly reduce seed and pods rot, improving germination rates, and hamper yield losses, thus enhancing sustainability of soybean production.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org. Table S1: The primers used for amplification and identification of Fusarium and Colletotrichum species. Table S2: Reference sequences of rDNA ITS gene from NCBI used for the homology analysis of isolated fungal diversity. Table S3: Reference sequences of RPB2 and of EF1-α gene from Fusarium MLST, GenBank and FUSARIUM-ID used for the homology analysis of isolated Fusarium species. Table S4: The GenBank accession numbers of RPB2 and EF1-α of Fusarium species obtained from soybean pods. Table S5: Reference sequences of ITS, CHS, GADPH, ACT, CAL and of TUB2 gene from NCBI GenBank used for the homology analysis of isolated Colletotrichum species. Table S6: The gene bank accession numbers of CHS, ITS, GADPH, ACT, CAL and TUB2 gene from NCBI. Table S7: Seed pathogenicity of Fusarium species isolated from intercropped soybean pods. Table S8: Seeds pathogenicity of Colletotrichum species isolated from intercropped soybean pods. Figure S1: Seed symptoms after inoculation with the representative isolates of different Fusarium species from intercropped soybean pods. Figure S2: Seed symptoms after inoculation with the representative isolates of different Colletotrichum species from intercropped soybean pods.
Author Contributions
Conceptualization, M.M. and X.L.; methodology, M.M.; software, M.M. and F.Y; validation, M.M., X.L. and W.Z.; formal analysis, M.M. and T.Y; investigation, M.M and X.L.; resources, X.L.; data curation, M.M. and Y.L.; writing—original draft preparation, M.M and M.N.; writing—review and editing, X,L., Z.S. and M.N.; visualization, X.L., S.Y. and X.W.; supervision, X.L.; project administration, X.L.; funding acquisition, X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key R&D Program (2023YFD1401000), Key Research and Development Plan of Sichuan Province (23ZDYF3037), and Guangxi Key Research and Development Program (AB23026107).
Data Availability Statement
The gene sequence information of Fusarium and Colletotrichum isolates in this study are available in the database of NCBI.
Acknowledgments
We are thankful to Dr. Gulshan Irshad for his kind guidance during fungal isolation.
Conflicts of Interest
The authors declare no conflict of interest in this work. All forms of financial support are acknowledged in the contribution. This work does not involve any human participants or animals. All authors have offered the consent to submission.
References
- Loganathan, M.; Maruthasalam, S.; Shiu, L.Y.; Lien, W.C.; Hsu, W.H.; Lee, P.F.; Yu, C.W.; Lin, C.H. Regeneration of soybean (Glycine max L. Merrill) through direct somatic embryogenesis from the immature embryonic shoot tip. In Vitro Cellular & Developmental Biology-Plant 2010, 46, 265–273. [Google Scholar]
- Kumari, S.; Dambale, A.S.; Samantara, R.; Jincy, M.; Bains, G. Introduction, history, geographical distribution, importance, and uses of soybean (Glycine max L.). In Soybean Production Technology: Physiology, Production and Processing; Springer: 2025; pp. 1-17.
- Modgil, R.; Tanwar, B.; Goyal, A.; Kumar, V. Soybean (Glycine max). In Oilseeds: health attributes and food applications; Springer: 2020; pp. 1-46.
- Belewu, M.; Belewu, K. Comparative physico-chemical evaluation of tiger-nut, soybean and coconut milk sources. International Journal of Agriculture and Biology 2007, 5, e787. [Google Scholar]
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In Abiotic and biotic stresses in soybean production; Elsevier: 2016; pp. 1-26.
- Mishra, R.; Tripathi, M.; Sikarwar, R.; Singh, Y.; Tripathi, N. Soybean (Glycine max L. Merrill): A multipurpose legume shaping our world. Plant Cell Biotechnol. Mol. Biol 2024, 25, 17–37. [Google Scholar] [CrossRef]
- Hosseini, B.; Voegele, R.T.; Link, T.I. Diagnosis of soybean diseases caused by fungal and oomycete pathogens: Existing methods and new developments. Journal of Fungi 2023, 9, 587. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.; Li, H.; Yan, L.; Raza, M.A.; Gong, G.; Chen, H.; Yang, C.; Zhang, M.; Shang, J.; Liu, T. Characterization and pathogenicity of Fusarium species associated with soybean pods in maize/soybean strip intercropping. Pathogens 2019, 8, 245. [Google Scholar] [CrossRef]
- Arias, M.M.D.; Leandro, L.F.; Munkvold, G.P. Aggressiveness of Fusarium species and impact of root infection on growth and yield of soybeans. Phytopathology 2013, 103, 822–832. [Google Scholar] [CrossRef]
- Barros, G.G.; Zanon, M.S.A.; Chiotta, M.L.; Reynoso, M.M.; Scandiani, M.M.; Chulze, S.N. Pathogenicity of phylogenetic species in the Fusarium graminearum complex on soybean seedlings in Argentina. European Journal of Plant Pathology 2014, 138, 215–222. [Google Scholar] [CrossRef]
- Chang, X.; Li, H.; Naeem, M.; Wu, X.; Yong, T.; Song, C.; Liu, T.; Chen, W.; Yang, W. Diversity of the seedborne fungi and pathogenicity of Fusarium species associated with intercropped soybean. Pathogens 2020, 9, 531. [Google Scholar] [CrossRef]
- Zhang, J.; Xue, A.; Cober, E.; Morrison, M.; Zhang, H.; Zhang, S.; Gregorich, E. Prevalence, pathogenicity and cultivar resistance of Fusarium and Rhizoctonia species causing soybean root rot. Canadian Journal of Plant Science 2013, 93, 221–236. [Google Scholar] [CrossRef]
- Pedrozo, R.; Little, C.R. Fusarium verticillioides inoculum potential influences soybean seed quality. European Journal of Plant Pathology 2017, 148, 749–754. [Google Scholar] [CrossRef]
- Pioli, R.; Mozzoni, L.; Morandi, E. First report of pathogenic association between Fusarium graminearum and soybean. Plant Disease 2004, 88, 220–220. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. The Fusarium laboratory manual; John Wiley & Sons: 2008.
- Chiotta, M.L.; Alaniz Zanon, M.S.; Palazzini, J.M.; Scandiani, M.M.; Formento, Á.N.; Barros, G.G.; Chulze, S.N. Pathogenicity of Fusarium graminearum and F. meridionale on soybean pod blight and trichothecene accumulation. Plant Pathology 2016, 65, 1492–1497. [Google Scholar] [CrossRef]
- Olszewski, J.; Dzienis, G.; Okorski, A.; Goś, W.; Pszczółkowska, A. Fungal Colonization of the Anatomical Parts of Soybean Seeds Supplied with Different Nitrogen Rates and Inoculated with Bradyrhizobium japonicum. Agriculture 2025, 15, 857. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, Z.; Chen, G.; Cao, A.; Wang, Q.; Yan, D.; Fang, W.; Li, Y. Detection and identification methods and control techniques for crop seed diseases. Agriculture 2023, 13, 1786. [Google Scholar] [CrossRef]
- Mancini, V.; Murolo, S.; Romanazzi, G. Diagnostic methods for detecting fungal pathogens on vegetable seeds. Plant Pathology 2016, 65, 691–703. [Google Scholar] [CrossRef]
- Panwar, S.; Duggirala, K.S.; Yadav, P.; Debnath, N.; Yadav, A.K.; Kumar, A. Advanced diagnostic methods for identification of bacterial foodborne pathogens: Contemporary and upcoming challenges. Critical Reviews in Biotechnology 2023, 43, 982–1000. [Google Scholar] [CrossRef]
- Echarte, L.; Della Maggiora, A.; Cerrudo, D.; Gonzalez, V.; Abbate, P.; Cerrudo, A.; Sadras, V.; Calvino, P. Yield response to plant density of maize and sunflower intercropped with soybean. Field Crops Research 2011, 121, 423–429. [Google Scholar] [CrossRef]
- Abdelmagid, A.; Hafez, M.; Soliman, A.; Adam, L.R.; Daayf, F. First report of Fusarium sporotrichioides causing root rot of soybean in Canada and detection of the pathogen in host tissues by PCR. Canadian Journal of Plant Pathology 2021, 43, 527–536. [Google Scholar] [CrossRef]
- Chang, K.; Hwang, S.; Conner, R.; Ahmed, H.; Zhou, Q.; Turnbull, G.; Strelkov, S.; McLaren, D.; Gossen, B. First report of Fusarium proliferatum causing root rot in soybean (Glycine max L.) in Canada. Crop Protection 2015, 67, 52–58. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, N.; Chang, K.-F.; Hwang, S.-F.; Strelkov, S.E.; Conner, R.L.; McLaren, D.L.; Fu, H.; Harding, M.W.; Turnbull, G.D. Genetic diversity and aggressiveness of Fusarium species isolated from soybean in Alberta, Canada. Crop Protection 2018, 105, 49–58. [Google Scholar] [CrossRef]
- Chang, X.; Dai, H.; Wang, D.; Zhou, H.; He, W.; Fu, Y.; Ibrahim, F.; Zhou, Y.; Gong, G.; Shang, J. Identification of Fusarium species associated with soybean root rot in Sichuan Province, China. European Journal of Plant Pathology 2018, 151, 563–577. [Google Scholar] [CrossRef]
- Yang, X.; Feng, F. Ranges and diversity of soybean fungal diseases in North America. Phytopathology 2001, 91, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wei, X.; Zheng, T.; Gou, Y.N.; Wang, J.; Deng, J.X.; Li, M.J. Evaluation of pathogenic Fusarium spp. associated with soybean seed (Glycine max) in Hubei Province, China. Plant Disease 2022, 106, 3178–3186. [Google Scholar] [CrossRef] [PubMed]
- Abdelmagid1; 2, A.; Hafez1; 3, M.; Lawley, Y.; Adam, L.; Daayf, F. First report of Fusarium cerealis causing root rot on soybean. 2018.
- Ellis, M.; Arias, M.D.; Jimenez, D.C.; Munkvold, G.; Leandro, L. First report of Fusarium commune causing damping-off, seed rot, and seedling root rot on soybean (Glycine max) in the United States. Plant Disease 2013, 97, 284–284. [Google Scholar] [CrossRef]
- Olszak-Przybyś, H.; Korbecka-Glinka, G.; Patkowska, E. Identification and pathogenicity of Fusarium isolated from soybean in Poland. Pathogens 2023, 12, 1162. [Google Scholar] [CrossRef]
- Chang, X.; Naeem, M.; Li, H.; Yan, L.; Liu, T.; Liu, B.; Zhang, H.; Khaskheli, M.; Gong, G.; Zhang, M. First report of Fusarium asiaticum as a causal agent for seed decay of soybean (Glycine max) in Sichuan, China. Plant Disease 2020, 104, 1542–1542. [Google Scholar] [CrossRef]
- Chang, X.; Yan, L.; Naeem, M.; Khaskheli, M.I.; Zhang, H.; Gong, G.; Zhang, M.; Song, C.; Yang, W.; Liu, T. Maize/soybean relay strip intercropping reduces the occurrence of Fusarium root rot and changes the diversity of the pathogenic Fusarium species. Pathogens 2020, 9, 211. [Google Scholar] [CrossRef]
- Arias, M.D.; Munkvold, G.; Ellis, M.; Leandro, L. Distribution and frequency of Fusarium species associated with soybean roots in Iowa. Plant disease 2013, 97, 1557–1562. [Google Scholar] [CrossRef]
- Cannon, P.; Damm, U.; Johnston, P.; Weir, B. Colletotrichum: current status and future directions. Studies in mycology 2012, 73, 181–213. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 fungal pathogens in molecular plant pathology. Molecular plant pathology 2012, 13, 414–430. [Google Scholar] [CrossRef]
- da Silva, L.L.; Moreno, H.L.A.; Correia, H.L.N.; Santana, M.F.; de Queiroz, M.V. Colletotrichum: species complexes, lifestyle, and peculiarities of some sources of genetic variability. Applied microbiology and biotechnology 2020, 104, 1891–1904. [Google Scholar] [CrossRef]
- Sharma, S.; Gupta, G.; Ramteke, R. Colletotrichum truncatum [(Schw.) Andrus & WD Moore], the causal agent of anthracnose of soybean [Glycine max (L.) Merrill]—A Review. Soybean Res 2011, 9, 31–52. [Google Scholar]
- Yang, H.-C.; Hartman, G.L. Methods and evaluation of soybean genotypes for resistance to Colletotrichum truncatum. Plant disease 2015, 99, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Hartman, G.L.; Rupe, J.C.; Sikora, E.J.; Domier, L.L.; Davis, J.A.; Steffey, K.L. Compendium of soybean diseases and pests; American Phytopathological Society, St. Paul, MN: 2015.
- Cai, L.; Hyde, K.; Taylor, P.; Weir, B.; Waller, J.; Abang, M.; Zhang, J.; Yang, Y.; Phoulivong, S.; Liu, Z. A polyphasic approach for studying Colletotrichum. Fungal Diversity 2009, 39, 183–204. [Google Scholar]
- Damm, U.; Woudenberg, J.; Cannon, P.; Crous, P. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Diversity 2009, 39, 45. [Google Scholar]
- Hyde, K.; Cai, L.; Cannon, P.; Crouch, J.; Crous, P.; Damm, U.; Goodwin, P.; Chen, H.; Johnston, P.; Jones, E. Colletotrichum—names in current use. Fungal Diversity 2009, 39, 147–182. [Google Scholar]
- Yang, H.-C.; Haudenshield, J.; Hartman, G. First report of Colletotrichum chlorophyti causing soybean anthracnose. Plant Disease 2012, 96, 1699–1699. [Google Scholar] [CrossRef]
- Yang, H.-C.; Haudenshield, J.S.; Hartman, G.L. Colletotrichum incanum sp. nov., a curved-conidial species causing soybean anthracnose in USA. Mycologia 2014, 106, 32–42. [Google Scholar] [CrossRef]
- Khakimov, A.; Salakhutdinov, I.; Omolikov, A.; Utaganov, S. Traditional and current-prospective methods of agricultural plant diseases detection: A review. In Proceedings of the IOP Conference series: earth and environmental science, 2022; p. 012002.
- Dayarathne, M.C.; Mridha, A.U.; Wang, Y. Diagnosis of fungal plant pathogens using conventional and molecular approaches. In Diagnostics of plant diseases; IntechOpen: 2020.
- Da Lio, D.; Cobo-Díaz, J.F.; Masson, C.; Chalopin, M.; Kebe, D.; Giraud, M.; Verhaeghe, A.; Nodet, P.; Sarrocco, S.; Le Floch, G. Combined metabarcoding and multi-locus approach for genetic characterization of Colletotrichum species associated with common walnut (Juglans regia) anthracnose in France. Scientific reports 2018, 8, 10765. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating species boundaries in Colletotrichum. Fungal Diversity 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Fuentes-Aragón, D.; Guarnaccia, V.; Rebollar-Alviter, A.; Juárez-Vázquez, S.B.; Aguirre-Rayo, F.; Silva-Rojas, H.V. Multilocus identification and thiophanate-methyl sensitivity of Colletotrichum gloeosporioides species complex associated with fruit with symptoms and symptomless leaves of mango. Plant Pathology 2020, 69, 1125–1138. [Google Scholar] [CrossRef]
- Du JunBo, D.J.; Han TianFu, H.T.; Gai JunYi, G.J.; Yong TaiWen, Y.T.; Sun Xin, S.X.; Wang XiaoChun, W.X.; Yang Feng, Y.F.; Liu Jiang, L.J.; Shu Kai, S.K.; Liu WeiGuo, L.W. Maize-soybean strip intercropping: achieved a balance between high productivity and sustainability. 2018.
- Yang, F.; Wang, X.; Liao, D.; Lu, F.; Gao, R.; Liu, W.; Yong, T.; Wu, X.; Du, J.; Liu, J. Yield response to different planting geometries in maize–soybean relay strip intercropping systems. Agronomy Journal 2015, 107, 296–304. [Google Scholar] [CrossRef]
- Liu, J.; Deng, J.; Zhang, K.; Wu, H.; Yang, C.; Zhang, X.; Du, J.; Shu, K.; Yang, W. Pod mildew on soybeans can mitigate the damage to the seed arising from field mold at harvest time. Journal of agricultural and food chemistry 2016, 64, 9135–9142. [Google Scholar] [CrossRef] [PubMed]
- Doohan, F.; Brennan, J.; Cooke, B. Influence of climatic factors on Fusarium species pathogenic to cereals. Epidemiology of Mycotoxin Producing Fungi: Under the Aegis of COST Action 835 ‘Agriculturally Important Toxigenic Fungi 1998–2003’, EU project (QLK 1-CT-1998–01380) 2003, 755-768.
- Liu, Y.; Wu, D.; Liu, Q.; Zhang, S.; Tang, Y.; Jiang, G.; Li, S.; Ding, W. The sequevar distribution of Ralstonia solanacearum in tobacco-growing zones of China is structured by elevation. European Journal of Plant Pathology 2017, 147, 541–551. [Google Scholar] [CrossRef]
- Jiang, G.; Wei, Z.; Xu, J.; Chen, H.; Zhang, Y.; She, X.; Macho, A.P.; Ding, W.; Liao, B. Bacterial wilt in China: history, current status, and future perspectives. Frontiers in Plant Science 2017, 8, 1549. [Google Scholar] [CrossRef]
- Zhou, Y.; Gong, G.; Cui, Y.; Zhang, D.; Chang, X.; Hu, R.; Liu, N.; Sun, X. Identification of Botryosphaeriaceae species causing kiwifruit rot in Sichuan Province, China. Plant Disease 2015, 99, 699–708. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P. Phylogenetic diversity of insecticolous fusaria inferred from multilocus DNA sequence data and their molecular identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular biology and evolution 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Gao, X.; Wu, M.; Xu, R.; Wang, X.; Pan, R.; Kim, H.-J.; Liao, H. Root interactions in a maize/soybean intercropping system control soybean soil-borne disease, red crown rot. Plos one 2014, 9, e95031. [Google Scholar] [CrossRef]
- Naeem, M.; Munir, M.; Li, H.; Raza, M.A.; Song, C.; Wu, X.; Irshad, G.; Khalid, M.H.B.; Yang, W.; Chang, X. Transcriptional responses of Fusarium graminearum interacted with soybean to cause root rot. Journal of Fungi 2021, 7, 422. [Google Scholar] [CrossRef]
- Miedaner, T.; Bolduan, C.; Melchinger, A. Aggressiveness and mycotoxin production of eight isolates each of Fusarium graminearum and Fusarium verticillioides for ear rot on susceptible and resistant early maize inbred lines. European Journal of Plant Pathology 2010, 127, 113–123. [Google Scholar] [CrossRef]
- Chang, X.; Wei, D.; Zeng, Y.; Zhao, X.; Hu, Y.; Wu, X.; Song, C.; Gong, G.; Chen, H.; Yang, C. Maize-soybean relay strip intercropping reshapes the rhizosphere bacterial community and recruits beneficial bacteria to suppress Fusarium root rot of soybean. Frontiers in Microbiology 2022, 13, 1009689. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Chen, H.K.; Ni, H.-F. Identification and characterization of Colletotrichum species associated with mango anthracnose in Taiwan. European Journal of Plant Pathology 2020, 157, 1–15. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Gilardi, G.; Faedda, R.; Schena, L.; Pane, A.; Garibaldi, A.; Gullino, M.L. Characterization of Colletotrichum ocimi population associated with black spot of sweet basil (Ocimum basilicum) in Northern Italy. Plants 2020, 9, 654. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Gilardi, G.; Martino, I.; Garibaldi, A.; Gullino, M.L. Species diversity in Colletotrichum causing anthracnose of aromatic and ornamental Lamiaceae in Italy. Agronomy 2019, 9, 613. [Google Scholar] [CrossRef]
- Liu, F.; Damm, U.; Cai, L.; Crous, P.W. Species of the Colletotrichum gloeosporioides complex associated with anthracnose diseases of Proteaceae. Fungal Diversity 2013, 61, 89–105. [Google Scholar] [CrossRef]
- Summerell, B.A. Resolving Fusarium: Current status of the genus. Annual review of phytopathology 2019, 57, 323–339. [Google Scholar] [CrossRef]
- O’Donnell, K.; Whitaker, B.K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Kim, H.S.; McCormick, S.P.; Busman, M.; Aoki, T. DNA sequence-based identification of Fusarium: A work in progress. Plant disease 2022, 106, 1597–1609. [Google Scholar] [CrossRef]
- Neergaard, P.; Neergaard, P. Management of Seed Storage. Seed Pathology: Volume I 1977, 574-594.
- Killebrew, J.; Roy, K.; Lawrence, G.; McLean, K.; Hodges, H. Greenhouse and field evaluation of Fusarium solani pathogenicity to soybean seedlings. 1988.
- Feng, J.; Hwang, R.; Chang, K.; Hwang, S.; Strelkov, S.; Gossen, B.; Conner, R.; Turnbull, G. Genetic variation in Fusarium avenaceum causing root rot on field pea. Plant Pathology 2010, 59, 845–852. [Google Scholar] [CrossRef]
- van Diepeningen, A.D.; Brankovics, B.; Iltes, J.; Van der Lee, T.A.; Waalwijk, C. Diagnosis of Fusarium infections: approaches to identification by the clinical mycology laboratory. Current fungal infection reports 2015, 9, 135–143. [Google Scholar] [CrossRef]
Figure 1.
The obtained mycoflora infecting soybean pods based on morphological features and sequence analysis of rDNA ITS fragments. (A) Colonial morphology of fungal isolates, (B) Phylogenetic tree of fungal isolates constructed using rDNA ITS. (C) The isolation frequency of fungal isolates associated with intercropped soybean pods. The colonies were observed after 7 days of incubation on PDA. The phylogenetic tree including 132 isolates and 25 reference isolates, and an outgroup Monilochaetes infuscans was constructed by MEGA X, and branches showing values >70 was excluded. Bootstrap support values were calculated from 1000 replications. Monilochaetes infuscans was used as outgroup.
Figure 1.
The obtained mycoflora infecting soybean pods based on morphological features and sequence analysis of rDNA ITS fragments. (A) Colonial morphology of fungal isolates, (B) Phylogenetic tree of fungal isolates constructed using rDNA ITS. (C) The isolation frequency of fungal isolates associated with intercropped soybean pods. The colonies were observed after 7 days of incubation on PDA. The phylogenetic tree including 132 isolates and 25 reference isolates, and an outgroup Monilochaetes infuscans was constructed by MEGA X, and branches showing values >70 was excluded. Bootstrap support values were calculated from 1000 replications. Monilochaetes infuscans was used as outgroup.
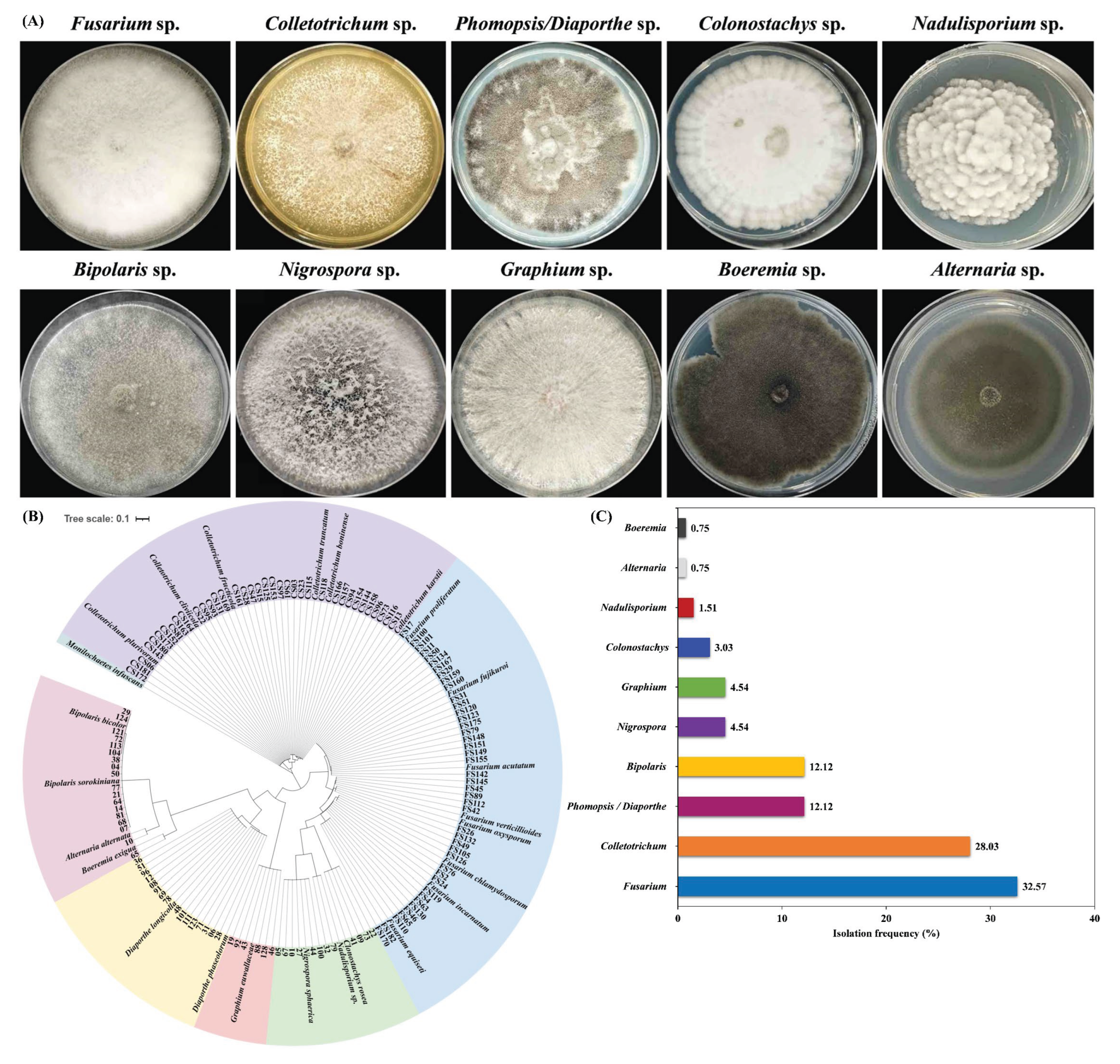
Figure 2.
Morphological and molecular identification of Fusarium isolates associated with soybean pods. (A) Characterization of colonies and conidium of Fusarium isolates. Typical colonies of Fusarium species were observed after 7 days, and conidium spores (down line) were visualized under microscopy after 10 days of growth on PDA. Scale bars are 20 μm. (B) Phylogenetic tree of Fusarium isolates based on rDNA ITS fragments. A maximum likelihood (ML) tree was constructed by MEGA X (Pennsylvania State University). Bootstrap support values were ≥50% from 1000 replications, which are shown at the nodes.
Figure 2.
Morphological and molecular identification of Fusarium isolates associated with soybean pods. (A) Characterization of colonies and conidium of Fusarium isolates. Typical colonies of Fusarium species were observed after 7 days, and conidium spores (down line) were visualized under microscopy after 10 days of growth on PDA. Scale bars are 20 μm. (B) Phylogenetic tree of Fusarium isolates based on rDNA ITS fragments. A maximum likelihood (ML) tree was constructed by MEGA X (Pennsylvania State University). Bootstrap support values were ≥50% from 1000 replications, which are shown at the nodes.
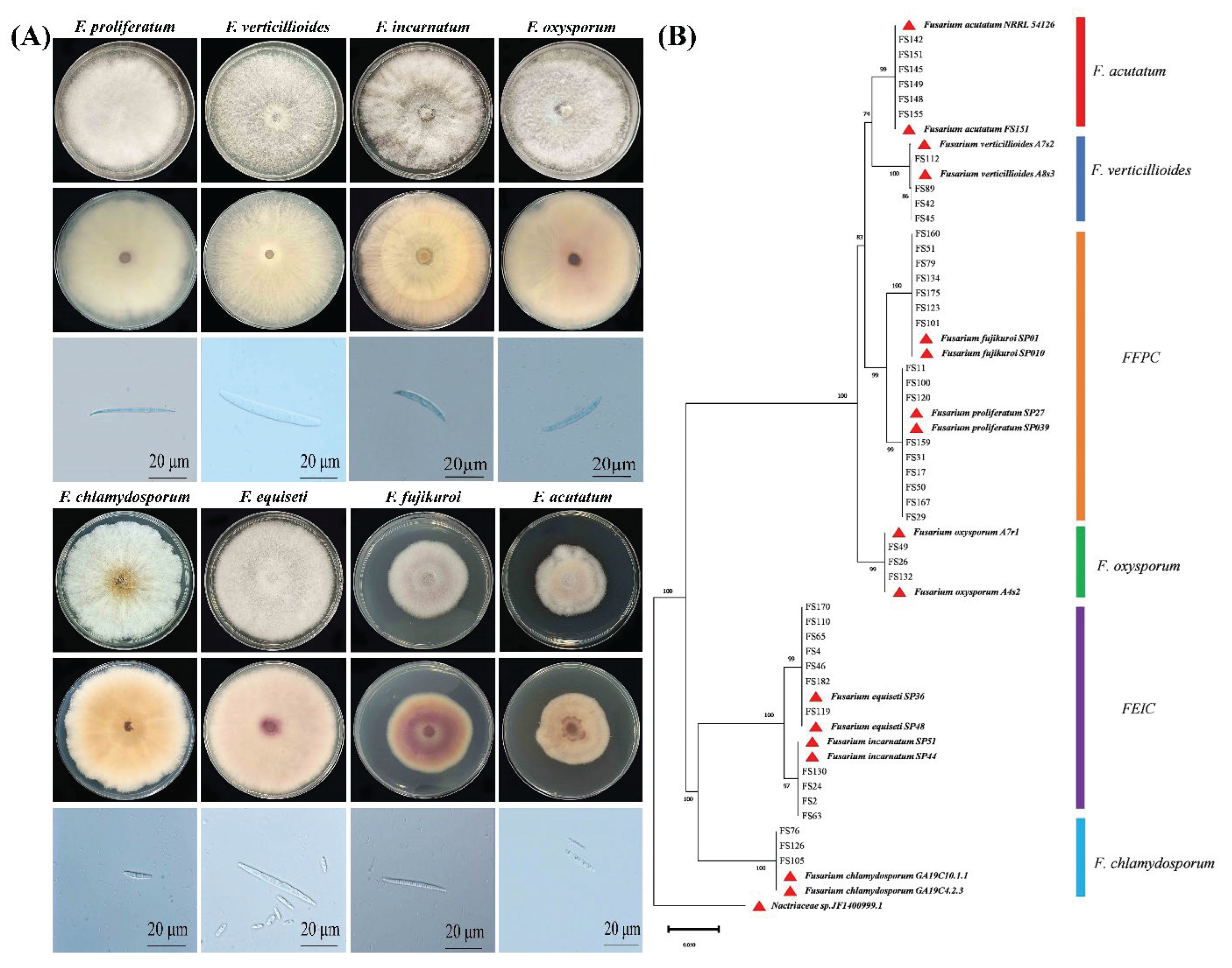
Figure 3.
Morphological and molecular identification of Colletotrichum isolates associated with soybean pods. (A) The morphological traits of Colletotrichum species isolated from soybean. Colonies of Colletotrichum species were examined after 7 days and spores after 15 days of incubation. (B) Phylogenetic tree of Colletotrichum isolates based on rDNA ITS, CHS, GAPDH, ACT, CAL and TUB2 genes. The tree was constructed using the ML method by MEGA X (Pennsylvania State University). Bootstrap support values were ≥50% from 1000 replications, which are shown at the nodes.
Figure 3.
Morphological and molecular identification of Colletotrichum isolates associated with soybean pods. (A) The morphological traits of Colletotrichum species isolated from soybean. Colonies of Colletotrichum species were examined after 7 days and spores after 15 days of incubation. (B) Phylogenetic tree of Colletotrichum isolates based on rDNA ITS, CHS, GAPDH, ACT, CAL and TUB2 genes. The tree was constructed using the ML method by MEGA X (Pennsylvania State University). Bootstrap support values were ≥50% from 1000 replications, which are shown at the nodes.
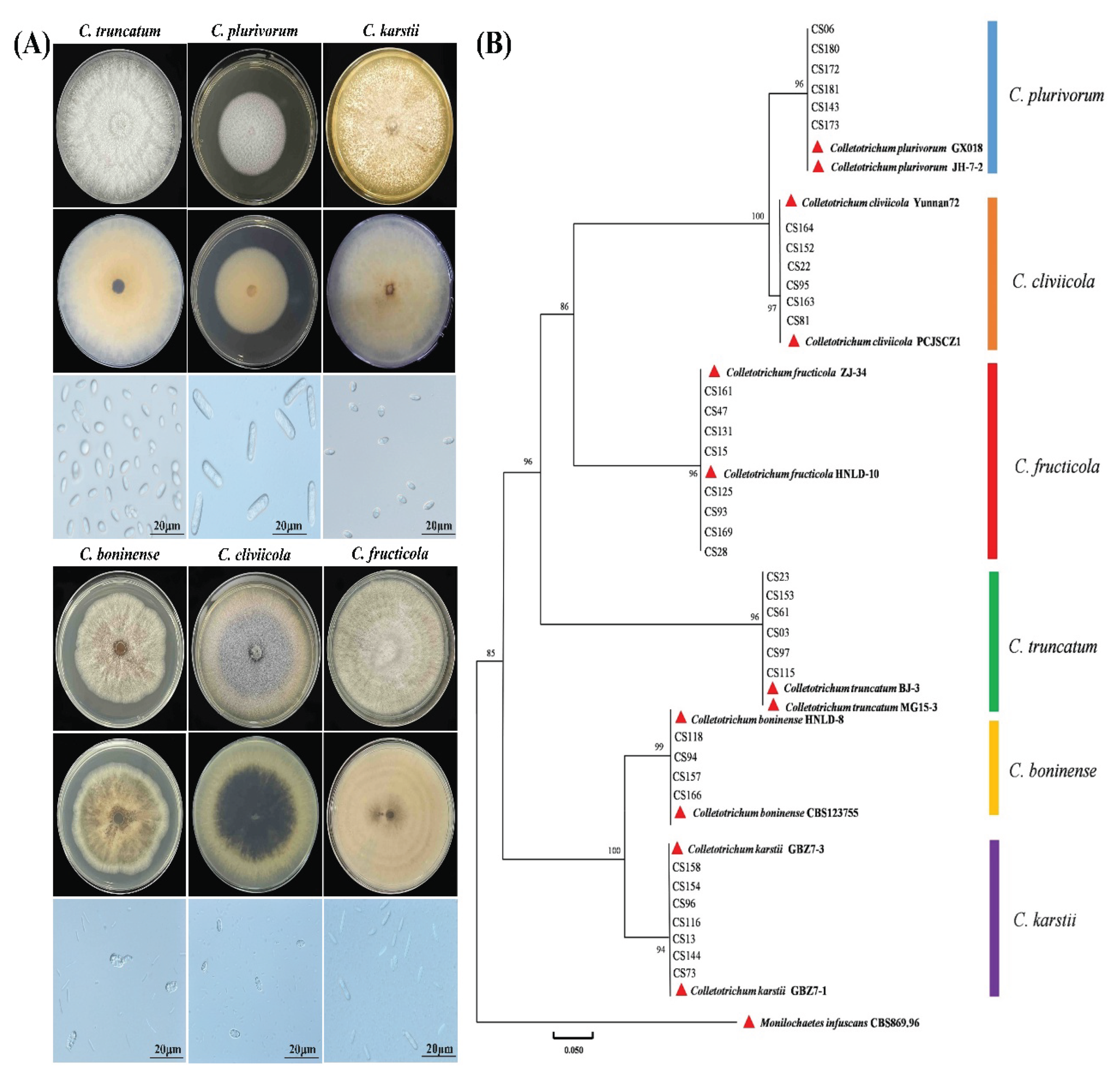
Figure 4.
The isolation frequency of Fusarium and Colletotrichum species from intercropped soybean pods. (A) Fusarium sp. and (B) Colletotrichum sp.
Figure 4.
The isolation frequency of Fusarium and Colletotrichum species from intercropped soybean pods. (A) Fusarium sp. and (B) Colletotrichum sp.
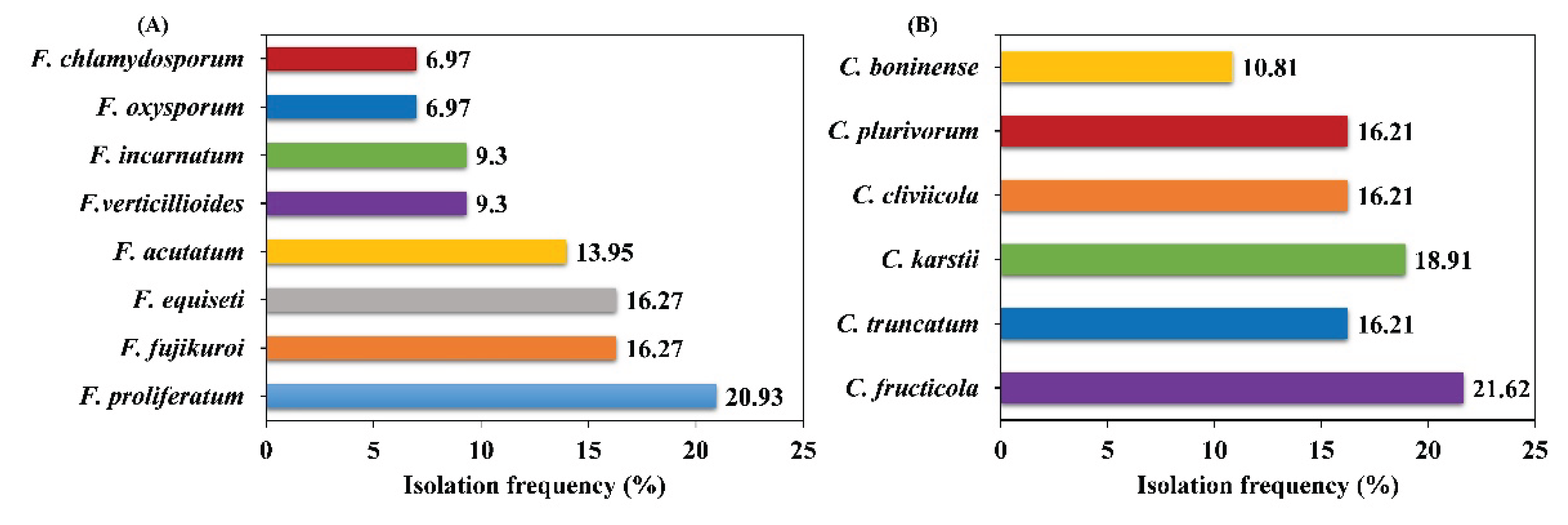
Figure 5.
The pathogenicity of different Fusarium species on soybean pods. Soybean pods were inoculated with the selected Fusarium isolates by a pod-soaking inoculation method at a final concentration of 1 × 105 spores per mL. The disease symptoms were observed after 7 days of post inoculation.
Figure 5.
The pathogenicity of different Fusarium species on soybean pods. Soybean pods were inoculated with the selected Fusarium isolates by a pod-soaking inoculation method at a final concentration of 1 × 105 spores per mL. The disease symptoms were observed after 7 days of post inoculation.
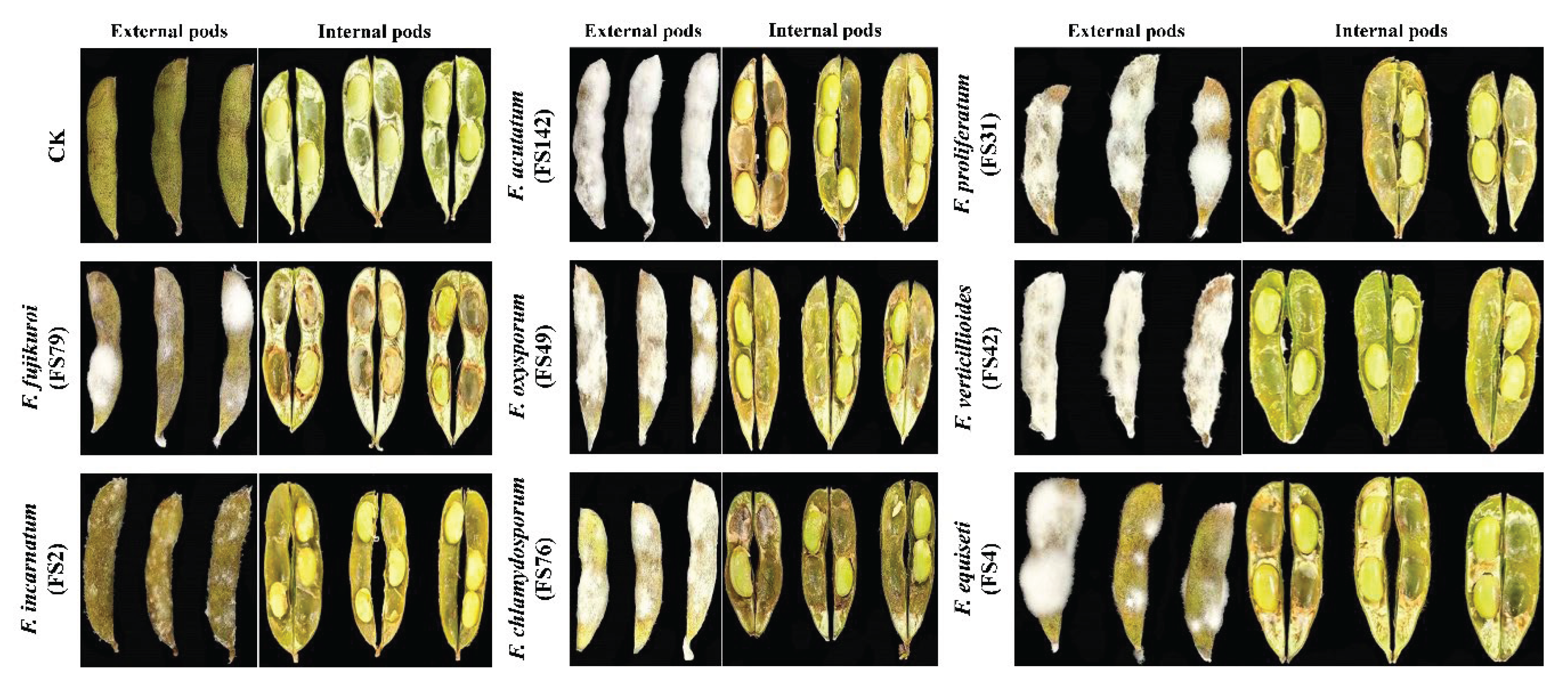
Figure 6.
The pathogenicity of different Colletotrichum species on soybean pods. Soybean pods were challenged by the chosen Colletotrichum isolates by using the pod-soaking inoculation method with the final concentration of 1 × 105 spores per mL. The disease symptoms were noted after 7 days of post inoculation.
Figure 6.
The pathogenicity of different Colletotrichum species on soybean pods. Soybean pods were challenged by the chosen Colletotrichum isolates by using the pod-soaking inoculation method with the final concentration of 1 × 105 spores per mL. The disease symptoms were noted after 7 days of post inoculation.
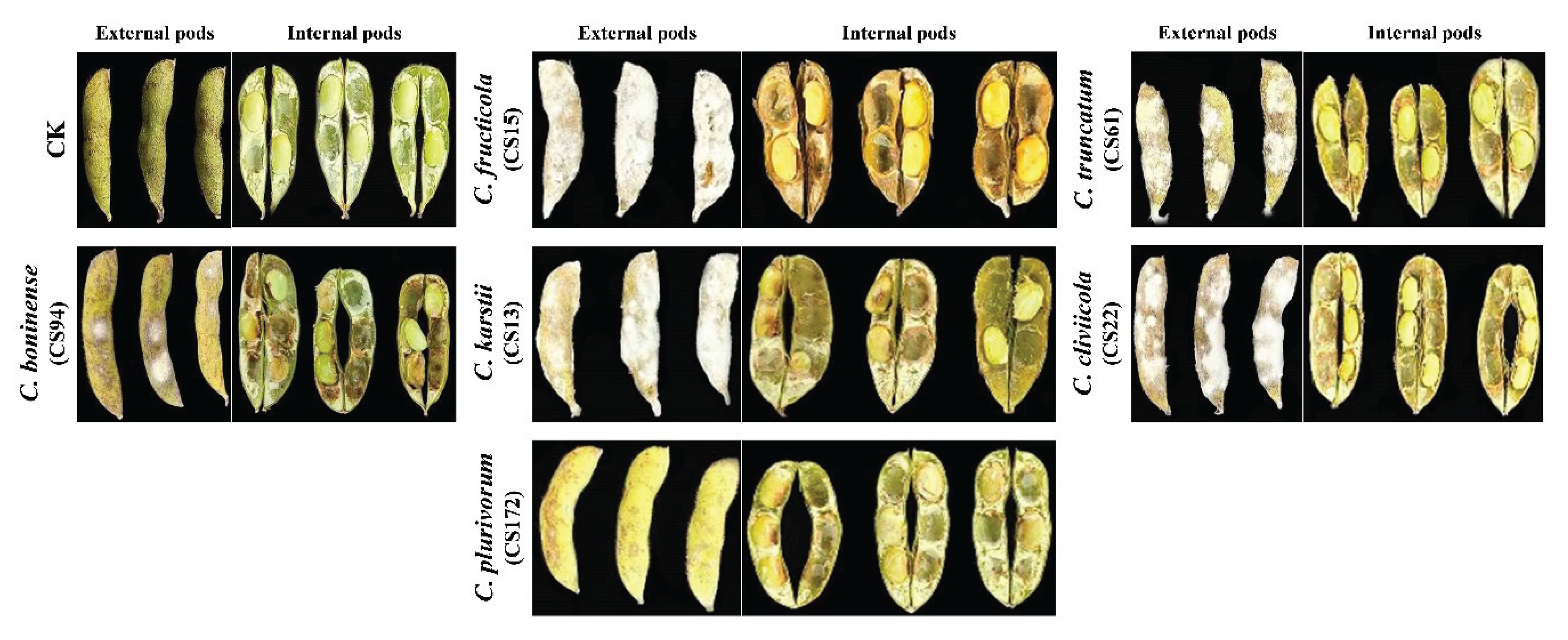

Table 1.
Morphological characters of Fusarium species cultured on PDA medium.
| Fusarium species | Macroconidia | Colony characterization | Growth rate (cm/day) | |||
|---|---|---|---|---|---|---|
| Shape | Width (μm) | Length (μm) | Septum | |||
| F. equiseti | Falcate | 3.10±0.02c, 3.02-3.7 |
39.25±1.81a, 38.23-45.98 |
3-5 | Pale grey color (front), ginger yellowish (back) | 4.88 ± 0.41b |
| F. incarnatum | Falcate | 3.98±0.44a, 5.67-2.72 |
36.98±3.63a, 45.55-36.62 |
3-4 | Pale grey color(front), yellowish color (back) | 5.32 ± 0.39a |
| F. verticillioides | Fusiform | 3.40±0.9b, 3.8-3.33 |
23.25±0.2b, 20.21-25.90 |
2-3 | pale grey colonies, reverse pale grey | 4.9 ± 0.3b |
| F. proliferatum | Falcate, fusiform | 3.60±1.12b, 5.41-2.96 |
39.12±6.54a, 48.56-32.66 |
3-4 | Pale grey color (front), pale grey (back) | 4.50 ± 0.03c |
| F. fujikuroi | Falcate | 2.42±0.46e, 3.12-2.28 |
39.92±1.98a, 43.82-38.94 |
3.-5 | Pale grey color (front), pale yellowish color (back) | 4.76 ± 0.32c |
| F. oxysporum | Falcate, with a foot spore | 3.1±0.16c, 2.31-4.82 |
26.9±1.6b, 28.62-22.23 |
3 | pale grey (front) pale purple on the back | 5.4 ± 0.3a |
| F. chlamydosporum | Falcate | 3.20±0.82c, 3.90-3.22 |
25.45±0.20b, 28.12-23.95 |
2-3 | Brown, light pink (Front) purple (back) | 5.23 ± 0.01a |
| F. acutatum | Falcate | 2.79±1.62d, 3.20-1.98 |
23.24±0.8b, 24.56-20.86 |
3-5 | white grey (front) purple (back) |
4.39 ± 0.02c |
Note: All data are the average value from three independent replicates. Different lower-case letter in the identical column reveals a significant variation at the level of P > 0.05.
Table 2.
Morphological characters of Colletotrichum species cultured on PDA medium.
| Species name | Texture | Conidial shape | Conidia size | Growth rate (cm/day) |
|
|---|---|---|---|---|---|
| Length (μm) | Width (μm) | ||||
| C. truncatum | Cottony | Fusiform | 23.20±0.56a 24.65-16.22 |
5.56±0.35b 5.90-4.32 |
6.90 ± 0.12a |
| C. karstii | Cottony | Cylindrical | 15.5±0.20b 18.20-14.90 |
6.80±0.23a 8.56-5.52 |
5.63 ± 0.27b |
| C. cliviicola | Cottony | oval/ellipsoidal | 13.35±0.02c 14.22-12.86 |
3.62±0.06c 4.56-3.22 |
6.12 ± 0.25a |
| C. plurivorum | Cottony and white | Fusiform | 13.75±0.12c 15.78-12.66 |
3.4±0.02c 4.45-3.56 |
6.30 ± 0.09a |
| C. boninense | Medium brown | Cylindrical | 15.10±0.2b 16.20-14.75 |
5.30±0.45b 6.60-4.25 |
6.15±0.02a |
| C.fructicola | Greyish black | Fusiform | 12.90±0.32c 14.56-10.86 |
6.80±0.23a 8.56-5.52 |
6.4±0.60a |
Note: All data are the means of three independent replicates: Different lower-case letter in the identical column reveals a significant variation at the level of P > 0.05. n=50.
Table 3.
Pathogenicity of Fusarium species on soybean pods isolated from intercropped soybean pods.
| Isolates | Disease incidence (%) | PMC (%) | DSI (%) | Pod Weight (g) |
|---|---|---|---|---|
| Control (CK) | 0 | 0±0e | 0±0d | 1.89±0.04d |
| F. proliferatum (FS31) | 100 | 90.66±0.40a | 96.66±4.71a | 1.79±0.31c |
| F. proliferatum (FS120) | 100 | 89.66±0.47a | 100±0a | 1.71±0.05c |
| F. proliferatum (FS167) | 100 | 78.33±2.35a | 93.33±4.71a | 1.74±0.01c |
| F. fujikuroi (FS79) | 100 | 43.33±2.35b | 66.66±11.78b | 2.48±0.27b |
| F. fujikuroi (FS101) | 100 | 46.66±4.71b | 75±0ab | 2.37±0.24b |
| F. fujikuroi (FS123) | 100 | 23.33±2.35c | 58.33±11.78b | 2.71±0.09b |
| F. equiseti (FS4) | 100 | 88.33±2.35a | 75±11.78b | 2.21±0.07b |
| F. equiseti (FS65) | 100 | 41.66±2.35b | 50±11.78b | 2.19±0.02b |
| F. equiseti (FS170) | 100 | 21.66±2.35c | 50±20.41b | 2.13±0.03b |
| F. acutatum (FS142) | 100 | 100±0a | 100±0a | 2.64±0.09b |
| F. acutatum (FS151) | 100 | 95±4.08a | 100±0a | 2.47±0.31b |
| F. acutatum (FS155) | 100 | 98.33±2.35a | 100±0a | 2.57±0.12b |
| F. verticillioides (FS42) | 100 | 98.33±2.35a | 100±0a | 2.29±0.12b |
| F. verticillioides (FS89) | 100 | 100±0a | 100±0a | 2.25±0.04b |
| F. verticillioides (FS112) | 100 | 100±0a | 100±0a | 2.29±0.06c |
| F. incarnatum (FS2) | 100 | 26.66±2.35c | 33.33±11.78c | 3.34±0.19a |
| F. incarnatum (FS24) | 100 | 20±4.08dc | 25±0c | 3.29±0.05a |
| F. incarnatum (FS130) | 100 | 18.33±6.23d | 33.33±11.78c | 3.25±0.06a |
| F. oxysporum (FS49) | 100 | 55±4.08b | 83.33±11.78a | 1.84±0.27c |
| F. oxysporum (FS26) | 100 | 45±4.08b | 66.66±11.78b | 1.86±0.04c |
| F. oxysporum (FS132) | 100 | 50±4.08b | 58.33±11.78b | 1.89±0.16c |
| F. chlamydosporum (FS76) | 100 | 18.33±2.35d | 75±0ab | 1.68±0.16c |
| F. chlamydosporum (FS105) | 100 | 23.33±6.23c | 50±20.41b | 1.57±0.05c |
| F. chlamydosporum (FS126) | 100 | 16.66±2.35d | 66.66±11.78b | 1.69±0.08c |
Note: The data are the average value from three independent replicates. Lowercase in the same column indicate significant difference. Significant difference was analyzed using Duncan’s multiple range assay at the level of P > 0.05.
Table 4.
Pods pathogenicity of Colletotrichum species isolated from intercropped soybean pods.
| Isolates | Disease incidence (%) | PMC (%) | DSI (%) | Pod weight (g) |
|---|---|---|---|---|
| Control (CK) | 0 | 0±0d | 0±0e | 1.86±0.040d |
| C. fructicola (CS15) | 100 | 90±4.08a | 100±0a | 2.51±0.04b |
| C. fructicola (CS93) | 100 | 86.66±6.23a | 91.66±11.78a | 2.52±0.02b |
| C. fructicola (CS169) | 100 | 76.66±2.35ab | 100±0a | 2.51±0.01b |
| C. truncatum (CS03) | 100 | 78.33±4.71ab | 75±0b | 2.09±0.02c |
| C. truncatum (CS61) | 100 | 80±4.08a | 83.33±11.78b | 2.11±0.01c |
| C. truncatum (CS153) | 100 | 81.66±2.35a | 66.66±11.7b | 2.16±0.08c |
| C. karstii (CS13) | 100 | 91.66±2.35a | 83.33±11.78b | 2.67±0.05b |
| C. karstii (CS 96) | 100 | 86.66±2.35a | 75±0b | 2.47±0.08b |
| C. karstii (CS 158) | 100 | 80±7.07a | 75±0b | 2.67±0.03b |
| C. cliviicola (CS 22) | 100 | 81.66±2.35a | 66.66±11.78c | 2.13±0.12c |
| C. cliviicola (CS95) | 100 | 76.66±2.35ab | 58.33±11.78c | 2.21±0.08c |
| C. cliviicola (CS 164) | 100 | 65±4.0b | 50±20.41c | 2.26±0.09c |
| C. plurivorum (CS06) | 100 | 10±4.08c | 25±0d | 2.60±0.02b |
| C. plurivorum (CS172) | 100 | 8.33±2.35c | 33.33±11.78d | 2.59±0.01b |
| C. plurivorum (CS180) | 100 | 6.66±2.35c | 25±20.41d | 2.61±0.01b |
| C. boninense (CS94) | 100 | 10±4.08c | 33.33±11.78d | 2.96±0.01a |
| C. boninense (CS118) | 100 | 11.66±6.23c | 25±0d | 2.98±0.02a |
| C. boninense (CS166) | 100 | 10±4.08c | 25±0d | 2.95±0.03a |
Note: The data are the mean value from three independent replicates. Lowercase in the same column indicate significant difference. Significant difference was calculated by applying Duncan’s multiple range assay at the level of P > 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



