
Submitted:
07 August 2025
Posted:
08 August 2025
You are already at the latest version
Abstract
A key aspect of climate change’s impact on organisms lies in understanding their ability to adapt to shifting and stressful environmental conditions. Insects, such as parasitoid wasps are particularly vulnerable due to limited heat tolerance. Adaptive strategies during mass rearing may enhance the efficacy and resilience of commercially reared biocontrol agents. This study assessed the effects of constant and fluctuating temperature regimens across four generations of mass-reared aphid parasitoids, examining their fitness traits and parasitism success under three thermal environments: colder [10°C], standard [20°C], and heat stress [28°C]. Parasitoids reared under fluctuating temperatures [day/night: 25°C /17°C] showed increased parasitism, but reduced progeny survival compared to those reared at a constant temperature [20°C]. Fluctuating regimens encouraged greater parasitism under heat stress, whereas constant regimens yielded intermediate parasitism across thermal environments, reflecting an evolutionary pattern consistent with the specialist-generalist trade-off. These findings underscore the value of evolutionary responses in developing adaptive rearing strategies that more accurately simulate field conditions, improving the performance of augmentative biological control under climate stress. Future research involving diverse fluctuating temperature regimes should deepen our understanding of trait trade-offs, such as survival and fecundity, and aid in identifying optimal thermal profiles to maximize resilience and efficacy in parasitoid wasps.
Keywords:
biological control
; parasitism
; natural enemy insects
; aphid parasitoids
; climate change
1. Introduction
The impacts of climate change are anticipated to profoundly disturb the interactions between natural enemies and insect pests, thereby affecting the efficacy of biocontrol programs, which in turn exacerbates the challenges that climate change poses to agricultural systems [1,2,3]. Biological control (natural and facilitated) is a crucial ecosystem service that entails reducing pest populations through natural control agents, such as parasitoids, predators, and pathogens, within either unmanaged or managed ecosystems [4,5,6]. Their environmental advantages have significantly accelerated the market demand for controllers in pest control programs in sustainable agricultural systems [6,7,8]. Although the magnitude of the consequences of climatic warming on biological control mediated by natural enemies is still less understood, these can involve a reduction in the effectiveness of biological control agents in the agroecosystem [9,10]. Furthermore, predicted climate change could exclude natural enemies from geographic areas where they currently persist [1]. Insects are particularly susceptible to climate change because most physiological processes are temperature dependent [11]. Environmental stresses such as increased temperature and increased climatic variability impact insect populations, affecting survival, development, reproduction, behavior, and phenology, and changing the abundance, distribution patterns, and seasonal timing of trophic interactions [12,13,14]. Parasitoid wasps (hymenopteran parasitoids) are important beneficial insects that play a critical ecological role in regulating other insect pests. They are highly specialized and synchronized with their herbivorous insect hosts, in which parasitoid eggs and larvae are developed, [15]. Both parasitoids and their insect hosts are strongly affected by environmental temperatures, but they respond differently to changes in temperature [1,16,17,18]. Several studies have shown differences in the thermal sensitivity between insect hosts and parasitoids, which could be common in nature, with parasitoid species severely threatened under increasing global temperatures as they often present a lower heat tolerance than their insect hosts [16,19,20,21,22]. For instance, the parasitoid Diadegma semiclausum could be negatively affected by increasing global temperatures due to the proximity of the optimal temperature to the critical thermal maximum in parasitoids as well as their limited plasticity in response to high temperatures than for their host Plutella xylostella, a global pest of Brassica crops [1]. Projections for climate change scenarios predict an overall increase in mean temperature and daily and seasonal variations in temperature, including rapid thermal changes such as increased incidence of high and extreme temperatures (e.g., heat waves) [23,24]. In this context, one major matter surrounding climate impacts on organisms is understanding how they respond to environmental stress and their ability to adapt to new changing environmental conditions [25,26,27]. In the short time, rapid adaptation in insects to changing climates is mainly mediated by plastic responses to thermal conditions such as seasonal strategies to overwinter (e.g., diapause) and thermal acclimation [2,28], which allow them to increase resistance to varying environmental conditions [29]. Plasticity is a major mechanism of phenotypic change and could be a critical strategy that allows individuals to cope with stresses arising under rapid environmental change [23,30], including acclimatization to environmental conditions within a generation (e.g., developmental acclimation and hardening) or across two or three generations (e.g., transgenerational and maternal effects) [28]. There is evidence of how prior exposition to thermal conditions during development can enhance thermotolerance in both host insects and parasitoid species within a single generation [31]. However, the responses observed are species-specific and can be strongly influenced by the thermal regimes experienced. For instance, studies on developmental acclimation in parasitoids of aphids have shown low support for beneficial acclimation [within a generation] to heat stress [32]. On the scale of multiple generations, intraspecific variation for thermal performance-related traits may evolve in response to the thermal environment by natural selection. Climate change is causing increases in both the mean and the variance of environmental temperature, each of which may act as a selective force on different traits of organisms [29,33,34]. For instance, the egg parasitoid Telenomus podisi used for controlling the brown stink bug (Euschistus heros) exposed to a fluctuating temperature regimen for four generations can positively influence fitness-related traits and costs of production compared with a constant temperature regimen [35]. Furthermore, it is necessary to determine whether the effect of rearing thermal conditions (mean /variance) shows patterns consistent with thermal adaptation. The thermal environment may act as an agent of selection for traits in populations that develop under contrasting thermal conditions. By examining the mean fitness of different populations or demes across a range of experimental habitats, it is possible to study the role of specific environmental factors as divergent selection agents. This approach enables a comparative analysis of fitness advantages in each population relative to its local environmental context, thereby elucidating the interaction patterns between demes and habitats in terms of fitness outcomes [36,37]. Understanding the potential of natural enemies to respond and adapt to changing environmental conditions is crucial to implementing climate adaptation measures in biological control programs. Despite the growing economic and environmental importance of managed biological control, there is still a knowledge gap in the relative importance of evolutionary responses to environmental factors on the performance of commercially reared agents. Augmentative biological control involves mass-producing natural enemies prior to their release to suppress pests in specific crops, where these lab-reared populations can be maintained over multiple generations under standard conditions, which provides an opportunity to develop and optimize biological control agents in controlled rearing environments, considering different thermal conditions that they may encounter after release. Endoparasitoid braconid wasps from the subfamily Aphidiinae (Hymenoptera: Braconidae) are an important group of natural enemies of sap-sucking aphid species (Hemiptera: Aphididae) [7]. They are used for commercial augmentative biocontrol in greenhouses and open crops to control agricultural aphid pests, which constitute important virus-transmitting pests on economically important crops [38]. These solitary parasitoid species present a high host specificity; a relatively short generation time and are easily breeding in the laboratory [15; 39]. Parasitoids of aphids lay an egg(s) (commonly a single egg) inside the body of aphid hosts and then develop through a larval stage to become a pupa where the body of the aphid is completely consumed, forming a called “mummy” (pupal stage of the parasitoid within the cuticle of its dead aphid host) from which only one parasitoid could emerge, adults can survive up to 20 days after emerging [40,41]. Here, we studied the generalist parasitoid Aphidius platensis Brethes [Hymenoptera: Braconidae: Aphidiinae] which can be found attacking important aphid pest species such as the peach potato, Myzus persicae, the cabbage aphid, Brevicoryne brassicae, Macrosiphum euphorbiae and Aulacorthum solani on diverse vegetables (cruciferous, lettuce, tomato, onion, marrow), cereal and horticultural crops [42,43,44]. The bird cherry-oat aphid, Rhopalosiphum padi Linnaeus, is an important winter host for A. platensis [45]. In addition, a recent study showed an absence of a diapause strategy in A. platensis populations studied from different aphid species and different climatic areas, indicating that individuals are active during the winter season [46,47]. This study first assessed the effect of temperature-rearing regimens (constant and fluctuating temperature regimens) on fitness-related traits during the fourth generation of mass-rearing in the aphid parasitoid A. platensis. We hypothesized that thermal environments during mass-rearing can influence the thermal adaptation of natural enemies; parasitoids reared under fluctuating environments can perform better under a range of thermal conditions than those reared under constant conditions. Then, it studied the effect of temperature regimens on the response of female parasitoids controlling aphid hosts across different temperature environments (colder-standard-heat stress conditions) to assess the role of environmental temperature underpinning the adaptation and optimization of mass-reared control agents for the improvement of augmentative biological control, especially in the context of climate change for which an increase in the mean and variance of temperature is expected [48].
2. Materials and Methods
Rearing Under Fluctuating and Constant Temperature Regimes
A single founder parasitoid population of the parasitoid A. platensis was established in the laboratory on its natal host, the aphid R. padi, in oat plants (aphid-plant systems) by collecting natural parasitoid populations from the field. Aphid parasitoids were sampled by collecting live aphids R. padi from oat fields and rearing under controlled laboratory conditions on oat (20°C, 60-75% RH, and photoperiod of 16:8 LD (Light: Dark, hours)) until the appearance of mummies (i.e., aphid exoskeletons containing the parasitoid pupae). To achieve high genetic and phenotypic variability (in traits under study), the founder parasitoid population was sampled in fields from two administrative regions among which agricultural production in Chile is mainly concentrated the central and southern Chile; Los Ríos and del Maule (39° to 35°S) (Mediterranean agroecosystem) [49], during spring growing season (November-December 2023). The emerged parasitoids (females and males) were maintained in a common environment that allowed random mating, after 48h the procedure consisted of allowing to single females to oviposit individually in closed plant-aphid systems (5 aphids). Following the development and emergence of parasitoid progeny in these systems, an ethanol sample of one female per system was taken for identification. Each female parasitoid individual was determined using taxonomic keys [42,43]. Then, only systems identified as A. platensis parasitoids were used to form the initial population. The founder population comprises a high number of founder parasitoids (>100 females previously allowed to mate) and it was kept under controlled conditions (20°C, 60-75% RH, 16:8 LD) by three generations for the remotion of maternal environmental effects before the establishment of alternative rearing regimes [28]. Then parasitoids were reared (mass-reared parasitoids) in two different temperature regimens carried out as follows: I) constant temperature regimen at 20°C (60-75% RH, 16:8 LD) (routinely used for mass-rearing biological control agents) and II) fluctuating temperature regimen at day 25°C and at night 17°C (60-75% RH, 16:8 LD) (approaching shifting conditions for field-releases). A constant temperature regimen was chosen based on the standard environmental condition, including the temperature of 20°C usually used to breed parasitoids on aphid cultures [50,51,52]. Fluctuating temperature conditions were chosen based on expected climatic indexes for increasing mean daily maximum and minimum temperatures for the growing season in the Mediterranean Chilean zone. To approach shifting conditions experienced by individuals for field releases. Further still, heat temperature does not represent a heat stress indicator (above 25°C) [53]. In each temperature regimen (constant or fluctuating), 6 experimental population cages were constituted for mass-reared parasitoid populations. Each one formed with ten parasitoid females with 48h-old mated that were randomly searched from a common emergence cage, containing female and male parasitoids that emerged from mummies from the previous generation and fed with water plus honey (2:1) [54,55,56]. Females were then reared on a standardized number of aphid hosts (100 aphids). A total of 100 adult aphids were added to the system 24 hours prior to the introduction of parasitoid females. Therefore, both the adult and nymphal stages of aphids can coexist in aphid-plant systems for parasitoid rearing. Both aphids and parasitoids were always kept in the same temperature regimen. Rearing environmental conditions for each regimen were carried out in a BIOBASE incubator, model BJPX-L200BK (Jinan City, China) (temperature volatility of ±0,5°C and temperature deviation of ±1°C). The system temperature was periodically controlled using an additional thermometer, with no variations being checked during the study period. After four generations of rearing, 30 female parasitoids with 48h-old mated were randomly chosen from temperature regimens for experimental assays. Each assay consisted of female single added-to-cage populations for choice and oviposition on 100 adult aphids for 24h in their respective temperature regimen (constant/fluctuating temperature). After 24h, females were removed alive from all experimental assays, which were keep in their temperature regimen until measurement of traits. The effect of temperature-rearing regimens on traits ecologically relevant for the performance of biological control agents in pest suppressing and mass rearing was experimentally studied by measuring the parasitism, estimated as the mean number of mummified aphids (mummies produced from standardized stung nymphs) [57]; survival of progeny as the number of parasitoids emerged from the number of mummies formed; developmental time estimated as the time from oviposition to the emergence of an adult parasitoid and, offspring sex ratio studied as the proportion of males in the progeny of a single female parasitoid. In hymenopteran parasitoids, females have control of the sex ratio by deciding the sex allocation through the fertilization of eggs or not [57,58]. The body size of progeny was measured as the hind-tibia length (mm) of female and males parasitoids, which is a standard measure to estimate the size of parasitoids [50], tooking pictures of the tibia using a digital camera Optikam PRO 5 (Optika, Italy) mounted on a stereo microscope Optika SZN-T ( × 45 ) and measures were taken using Optika vision Pro v2.7 software (Optika, Italy). Finally, the developmental growth rate of parasitoid progeny (females and males) in the different temperature regimens was estimated as the ratio between body size and total development time [59].
Effect of Rearing Temperature Regimen on the Response of Parasitoids Under Contrasting Thermal Conditions: A Reciprocal Transplant Experiment
To study the effect of mass-rearing conditions on the climatic pre-adaptability of natural enemies exposed to a thermal range, a reciprocal transplant experiment was carried out. The thermal adaptation pattern of parasitoids was evaluated by studying the performance of pest suppression in female parasitoids reared under different temperature regimens (constant and fluctuating regimens) when exposed to different temperatures during four generations of breeding. The temperature conditions included: (1) cold condition at 10°C (60-75% RH, 16:8 LD), (2) standard condition at 20°C (60-75% RH, 16:8 LD), and (3) stress heat condition at 28°C (60-75% RH, 16:8 LD). Contrasting conditions are based on an analysis of agro-climatic indices for the Chilean central valley. The cold condition represents the annual mean minimum temperature from different zones under study (Maule and Los Rios regions) (Araya-Osses et al. 2020). Under these conditions, Aphidius parasitoids from temperate climates could be induced into winter diapause, maintaining this state until spring (Tougeron et al. 2018). The standard condition represents the mean maximum temperature among geographical zones and is routinely used for mass-rearing of biological control agents [16,51,60]. The stress heat condition represents an agro-climatic index for summer, with heat stress temperatures above 25°C [53]. For experimental assays, female parasitoids were obtained from rearing cage populations; a plastic box (10 x 20 x 15 cm) with females and males for mating supplied with water and diluted honey (50%) for feeding, from the respective temperature regimen. A total of 21 female parasitoids 48-h old mated were randomly chosen from each different temperature regimen (constant and fluctuating temperature) were transferred to respective thermal conditions (cold, standard, or stress heat condition) with water and food for 24 hr. before assays. This period corresponds to a short exposure for acclimation to the temperatures tested with a duration of 24 h (hardening) [28]. Post-acclimation females were individually placed in the cage population (oat-aphid system) for choice and oviposition on 50 adult aphids for 24h to each temperature condition. Once 24 h had passed, each female was removed alive from the experimental assay, then each assay was transferred to a common thermal environment of 20°C until the measurement of the total number of mummies produced by parasitoid female in 24h in a thermal condition. In this way it is possible to focus the study on the parasitoid responses related to foraging behavior of females involving host choice and selection to oviposition by females, all ecologically relevant traits for the ability of parasitoids to suppress a pest population [15]. This experimental design has been used in similar studies on the effect of environmental temperature on parasitoid behavior [32].
Data Analysis
The effect of the rearing temperature regimen (constant or fluctuating) (fixed factor) was studied on the different variables measured in this study. The mean number of mummified aphids by female parasitoid was studied using a generalized linear mixed model (GLMM) with a negative binomial distribution implemented in the R package glmmTMB [61,62]. Smooth density curves of the number of mummified aphids were visualized using the Kernel Density Estimation (KDE) method implemented in Jamovi 2.6. in R [63,64]. Progeny survival was analyzed as the proportion of emerged and non-emerged (binary variable) in the parasitoid progeny using binary logistic regression with temperature regimen as predictor variable using Jamovi 2.6. in R [63,64]. Sex ratio of parasitoid progeny was analyzed using a GLMM with a binomial (logit) distribution for proportion data. The variables development time (days) and hind-tibia length (body size) were analyzed using GLMMs with a Gaussian distribution including both temperature-rearing and parasitoid sex as fixed factors. The body size variable was also transformed using Box-Cox transformation to correct variances with the ‘boxcox’ function from the MASS package in R [65,66]. The growth rate was analyzed using GLS model (Generalized Least Squares) for fitting within-group heteroscedasticity implemented in nlme package in R [67], the model included as fixed factors both temperature-rearing regimen and parasitoid sex. The linear relation between hind-tibia length and development time was determined through linear regression using Jamovi 2.6. in R [63]. The analysis of experimental assays to determine the effect of parasitoid populations’ rearing regimens on the response of parasitoid females on pest suppression (mean number of mummies produced by parasitoid females exposed to different thermal conditions, included the variables; rearing temperature regimen (constant or fluctuating) and thermal condition (stress, standard and cold condition) and its interaction in the GLMMs using negative binomial distribution. Post hoc comparisons were performed in the R package emmeans [68]. The significance of fixed effects was provided by using ‘Anova’ function of the car package [69]. Heteroscedasticity, normality, overdispersion, or zero-inflated data were checked in the models using the R package performance [70]. In addition, to correctly classify the distribution family from the models for our data, we used the function check distribution implemented in the R package performance [70].
3. Results
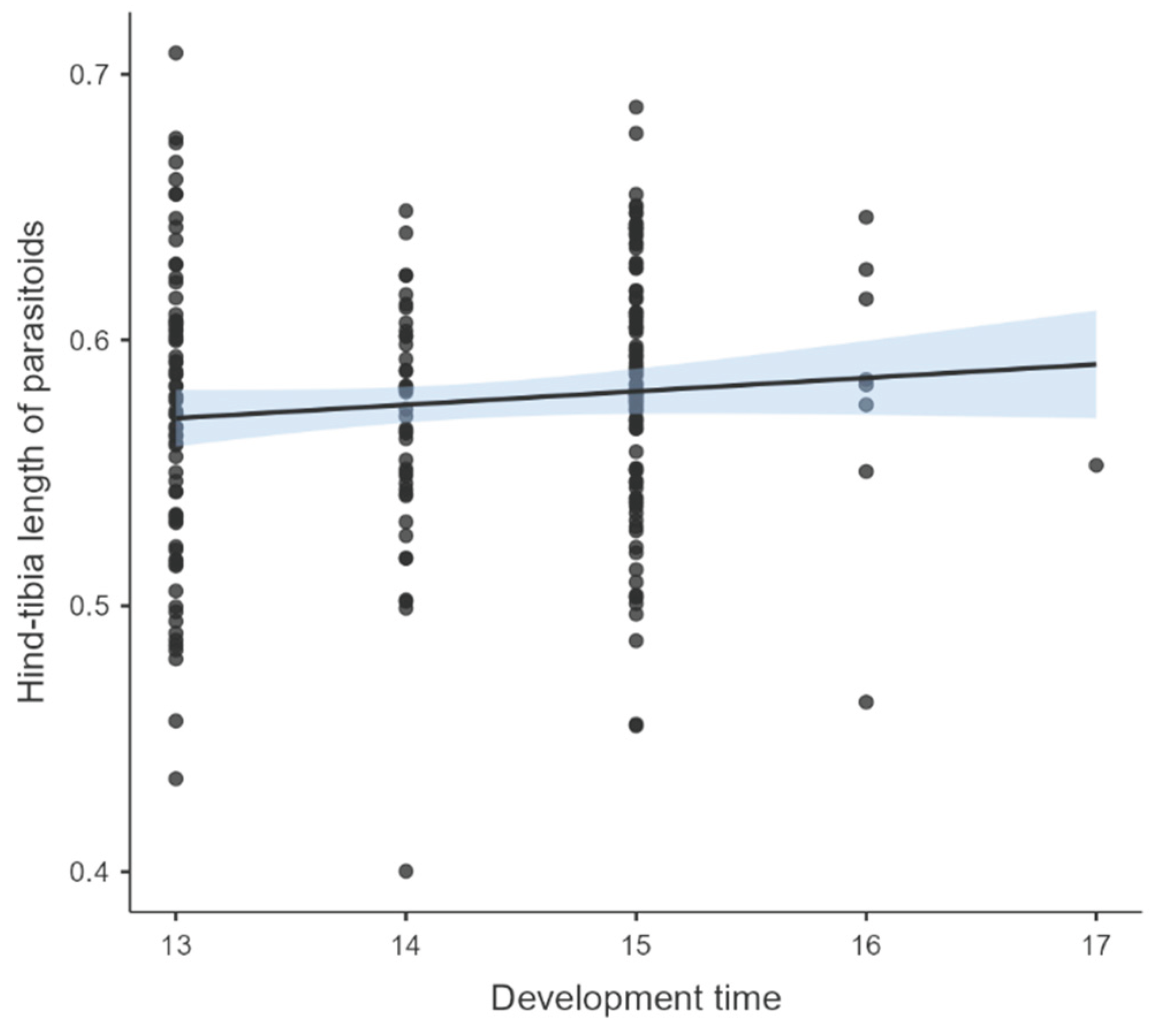
After four generations of parasitoid rearing, the effects of the temperature-rearing regimen on several fitness-related traits were studied in the parasitoid populations. The results shown a significant effect of temperature-rearing on the parasitism of female parasitoids; with a higher mean number of mummified aphids by female produced under fluctuating than in constant temperature regimen (Ꭓ2= 9.60; df = 1; P = 0.001) (Figures 1A and B). A significant effect of temperature-rearing regimen was found on the proportion of emerged parasitoids in the model of logistic regression (Ꭓ2= 13.3; df = 1; P < 0.001). From mummies formed (N=315), a higher proportion of emerging parasitoids in a constant than fluctuating temperature-rearing regimen was evidenced (OR= 0.361; P < 0.001) (Figure 1C). The mean sex ratio of parasitoid progeny did not show significant differences (Ꭓ2= 0.031; df = 1; P = 0.858) when rearing under fluctuating (mean 0.32 ± 0.08 SE) and constant regimen (mean 0.31 ± 0.07 SE), and this was female-biased (sex ratio < 0.5) in both regimens. The mean development time of progeny in mass-rearing parasitoid populations showed significant differences between temperature-rearing regimens (Ꭓ2= 129.62; df = 1; P < 2.2e-16), with a higher mean development time of progeny in the constant regimen than fluctuant regimen (Figure 2A). Similarly, the mean hind-tibia length of parasitoids was significantly affected by the temperature-rearing regimen (Ꭓ2 = 8.14; df= 1; P = 0.004), with more larger parasitoids in constant than fluctuating regimen (Figure 2B). Also, the body size was significantly influenced by parasitoid sex (Ꭓ2 = 5.76; df= 1; P = 0.016), observing larger sizes in females than males, but non-interaction effect between temperature regimen and sex was evidenced for this variable (Ꭓ2 = 0.06; df = 1; P = 0.800) (Figure 2B). The results did not show a relation between development time and body size of parasitoids (N=228) (r2 = 0.009; P = 0.151) (Appendix A). Temperature-rearing regimen showed a significant effect on growth rate of parasitoid progeny (Ꭓ2 = 13.93; df=1; P = 0.0001), which was higher in fluctuating than constant regimen, while non-significant effects were observed for the parasitoid sex (Ꭓ2 = 1.66; df=1; P = 0.197) nor the interaction between factors (Ꭓ2 = 0.86; df=1; P = 0.351) (Figure 2C).
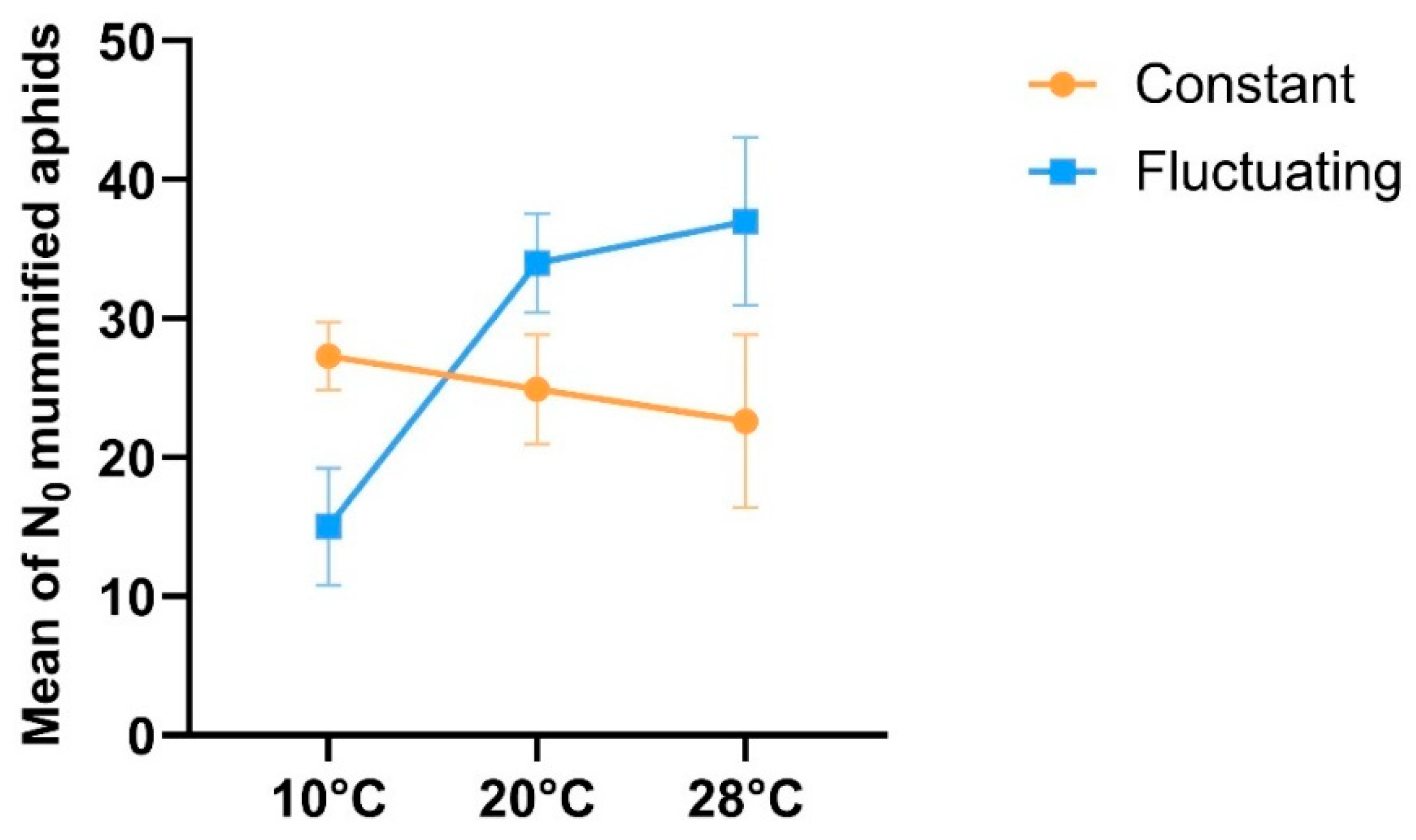
A significant interaction of temperature rearing regimen (fluctuating /constant regimens) in different tested thermal conditions (cold/standard/heat stress conditions) on the response of parasitism of parasitoid females was found at the four generations of rearing (temperature regimen x thermal condition interaction: Ꭓ 2= 12.86; df= 2; P = 0.0016), showing different in their mean response to the thermal conditions studied (Ꭓ2 = 9.21; df = 2; P = 0.009), while rearing regimen factor was not significant (Ꭓ2 = 0.004; df = 1; P = 0.947). The parasitism of parasitoid females in the heat stress temperature and standard condition was higher in comparison to the parasitism in cold conditions, whereas females in the constant temperature regimen showed similar intermediate parasitism level across different thermal conditions (Figure 3). The mean emergence rate of progeny for parasitoids did not show differences among different temperature regimens (Ꭓ2 = 1.21; df = 1; P = 0.270), thermal conditions (Ꭓ2 = 1.11; df = 2; P = 0.573) nor their interaction (temperature regimen x thermal condition) (Ꭓ2 = 1.66; df = 2; P = 0.434). The mean emergence rate, including all experimental assays, was 0.92 ± 0.007 (SE), and they were kept in a common standard temperature of 20°C.
4. Discussion
Environmental conditions, such as temperature, substantially impact the mass-rearing production of biocontrol agents [71,72]. Furthermore, these conditions can be optimized to match the selection pressures in the culture to those experienced in the field, including the changing climate [29,73]. After four generations of breeding parasitoid populations, the results showed an increased number of parasitized hosts when parasitoid wasps were reared under fluctuating temperatures in comparison with the constant temperature regimen (Figures 1A and B). Fecundity is a central fitness component and can thus be used as a proxy to assess the fitness consequences at different temperatures in parasitoids [74]. However, under fluctuating rearing conditions, parasitoids exhibited compromised survival (emergence rates). Specifically, a lower mean emergence rate (65%) was observed under the fluctuating rearing regimen compared to constant temperature-rearing conditions (85%). This suggests that the fluctuating regimen, particularly the high daily temperature of 25°C experienced during development, imposes strong selective pressure on parasitoid survival, a phenomenon also observed in other aphid parasitoid species [75]. Temperature can drive directional selection on trait variation within populations, leading to shifts in optimal trait values (e.g., thermal performance) over generations [76]. It can also impose evolutionary constraints, such as trade-offs in traits associated with fitness. For instance, positive environmental effects on certain traits may incur costs for others (allocation trade-offs), such as larger body size resulting in longer development times [59,77]. Our results indicated that parasitoids (both females and males) reared under constant temperatures exhibited larger body sizes, longer development times, along with lower growth rates compared to those reared under fluctuating temperatures. This aligns with the temperature-size rule [78], where individuals developing at lower temperatures achieve larger body sizes through a reduced growth rate, albeit at the cost of extended development times—a pattern widely documented in aphid parasitoids [59,75]. Larger females tend to exhibit greater longevity and higher egg loads, potentially enhancing their fecundity [52,59]. Conversely, shorter development times may facilitate more generations per year (voltinism), enabling rapid population growth and higher densities [59]. Such outcomes could improve the cost-effectiveness of mass-rearing programs and release strategies. Elevated temperature rearing can affect, to a greater degree, parasitoid development, decreasing larval survival, longevity, egg load, and body size and then affect their fecundity potential. This has been observed in related Aphidius parasitoids (Aphidius colemani Viereck and Aphidius rhopalosiphi De Stefani Perez) exposed to high temperatures [59,75]. These findings suggest limited capacity for plastic responses through acclimatization within a generation to adapt to heat stress, as high temperatures adversely impact multiple traits. For example, a study on A. colemani revealed limited support for the developmental acclimation hypothesis regarding heat tolerance in adults at 28°C, highlighting constraints on phenotypic plasticity [32].
Parasitoid fitness under temperature regimens is shaped by the thermal tolerance/niche of a population, as defined by the thermal performance curve (TPC), which quantifies performance or fitness across critical thermal limits [74]. Rearing temperatures can differentially affect parasitoid performance depending on whether they fall within the optimal range, where performance is maximized [79]. For several aphid parasitoid species (A. rhopalosiphi, Aphidius matricariae, and A. colemani), an optimal temperature has been reported near 20°C [52,75,80,81]. Our reciprocal transplant experiment revealed adaptive changes in response to thermal regimens experienced across parasitoid generations. Female parasitoids reared in a fluctuating regimen with high daily temperatures (at day: 25°C; at night: 17°C) showed increased parasitism under stress heat conditions (28°C) and under mild temperatures of the standard condition (20°C). While females from constant temperature regimen showed an intermediate parasitism across temperature range tested, including the cold condition at 10°C. Which was detrimental to the parasitoid performance from the fluctuating temperature regimen, suggesting a narrow performance breadth under fluctuating than constant regimen, consistent with generalist phenotype patterns (similar degree of performance to all habitats). This aligns with the conceptual model of thermal evolution, wherein rising mean temperatures favor genotypes with higher thermal optima but narrower performance breadths—a specialist-generalist trade-off [29]. The conceptual model of evolution in response to changes in environmental temperature suggests that with increases in mean environmental temperature, selection should favor genotypes that confer higher thermal optimum (maximal performance) for ecologically important activities at warmer temperatures, but as maximal performance increases, performance breadth (temperature range) should then decline as an indirect result of a specialist-generalist tradeoff [29]. For instance, adaptation to higher mean temperatures may lead to maladaptation to variability, as evidenced by reduced parasitism at 10°C by parasitoids from fluctuating regimens. In contrast, intermediate temperatures (e.g., constant 20°C) may foster generalist phenotypes with broader performance breadths [29,36]. This was reflected in females reared at 20°C, which displayed intermediate performance across thermal conditions, including cold stress, suggesting greater phenotypic plasticity. The standard condition of rearing to a temperature of 20°C (constant) used in this study could maintain a higher variation in the plasticity phenotypic within A. platensis populations, including a higher cold tolerance. A temperature of 20°C is considered the optimal condition for rearing within the thermal range of aphid parasitoids and their hosts, which potentially enhances the success of intermediate generalist phenotypes [29,36]. Natural populations of A. platensis exhibit high cold tolerance, remaining active during winter without diapause [46]. Such plasticity may enhance population persistence under climate variability. However, climate change is increasing exposure to extreme events, particularly heatwaves [23]. Thus, thermotolerance acquired through multigenerational exposure to warm regimens could optimize field performance of biocontrol agents. Besides, strong survival constraints under thermal stress may disproportionately shape the evolution of TPCs [74]. Trade-offs in resource allocation—e.g., between longevity and reproduction or size and development time—highlight constraints on plasticity and adaptation [31,59,77]. Understanding these trade-offs is critical for biological control efficacy, particularly in field releases. The observed trade-offs between parasitism and offspring survival under thermal environments need to be considered, as these may have complex effects on the actual biological control efficacy, especially when releasing parasitoids on a field scale. Future research should explore the physiological and molecular basis of these trade-offs and test more extreme or variable temperature regimen to better simulate natural conditions. Such efforts could refine rearing strategies and enhance the resilience of biocontrol agents under warming climate scenarios [82].
Author Contributions
“Conceptualization, F.Z.P. and B.L; methodology, F.Z.P., C.V. and M.V.; formal analysis, F.Z.P.; investigation, F.Z.P., C.V. and M.V.; writing—original draft preparation, F.Z.P.; writing—review and editing, B.L.; visualization, F.Z.P; supervision, F.Z.P.; project administration, F.Z.P.; funding acquisition, F.Z.P. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation.
Funding
This research was funded by Agencia Nacional de Investigación y Desarrollo, grant number 11230372 and The APC was funded by FONDECYT n° 11230372.
Institutional Review Board Statement
The study protocol was approved by the Institutional Biosecurity Committee of Universidad de Talca for studies involving animals.
Data Availability Statement
Data deposited in the Mendeley repository, doi: 10.17632/nk7xmgbzr7.1
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Figure A1.
Relationship Between Hind-Tibia Length of Parasitoids and Development Time in the Parasitoid Progeny Studied (r2 = 0.009; P = 0.151).
Figure A1.
Relationship Between Hind-Tibia Length of Parasitoids and Development Time in the Parasitoid Progeny Studied (r2 = 0.009; P = 0.151).
References
- Furlong, M.J., Zalucki, M.P. Climate change and biological control: the consequences of increasing temperatures on host–parasitoid interactions. Curr Opin Insect Sci. 2017; 20(1):39–44. [CrossRef]
- Tougeron, K., Brodeur, J., Le Lann, C., van Baaren, J. How climate change affects the seasonal ecology of insect parasitoids. Ecol Entomol. 2020; 45(2):167–181. [CrossRef]
- Monticelli, L.S., Bishop, J., Desneux, N., Gurr, G.M., Jaworski, C.C., McLean, A.H., Vanbergen, A.J. Multiple global change impacts on parasitism and biocontrol services in future agricultural landscapes. Adv Ecol Res. 2021; 65:245–304. [CrossRef]
- Barratt, B.I.P., Moran, V.C., Bigler, F., Van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl. 2018; 63(1):155–167. [CrossRef]
- Baker, B.P., Green, T.A., Loker, A.J. Biological control and integrated pest management in organic and conventional systems. Biol Control. 2020; 140:104095. [CrossRef]
- Stenberg, J.A. A conceptual framework for integrated pest management. Trends Plant Sci. 2017; 22(10):759–769. DOI: 10.1016/j.tplants.2017.06.010 .
- Van Lenteren, J.C., Bolckmans, K., Köhl, J., Ravensberg, W.J., Urbaneja, A. Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl. 2018; 63(1):39–59. [CrossRef]
- Patel, H.R., Mandaliya, V.B. Biocontrol agents in agriculture: patent landscape, market dynamics, and recommendations for sustainable farming. In: Mitra, D., de los Santos Villalobos, S., Rani, A., Guerra Sierra, B.E., Andjelković, S., editors. Bio-control Agents for Sustainable Agriculture. Singapore: Springer; 2025. [CrossRef]
- Romo, C.M., Tylianakis, J.M. Elevated temperature and drought interact to reduce parasitoid effectiveness in suppressing hosts. PLoS ONE. 2013; 8(3):e58136. [CrossRef]
- Wakil, W., Kavallieratos, N.G., Eleftheriadou, N., Ghazanfar, M.U., El-Shafie, H.A., Blankson, A., Harvey, J.A. Climate change consequences for insect pest management, sustainable agriculture and food security. Entomol Gen. 2025; 45(1):37–51. [CrossRef]
- Bale, J.S., Masters, G.J., Hodkinson, I.D., Awmack, C., Bezemer, T.M., Brown, V.K., et al. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Glob Change Biol. 2002; 8(1):1–16. [CrossRef]
- Choudhary, J.S., Kumari, M.K., Fand, B.B. Linking insect pest models with climate change scenarios to project against future risks of agricultural insect pests. CABI Rev. 2019; 2019:1–13. [CrossRef]
- Pardikes, N.A., Revilla, T.A., Lue, C.H., Thierry, M., Souto-Vilarós, D., Hrcek, J. Effects of phenological mismatch under warming are modified by community context. Glob Change Biol. 2022; 28(13):4013–4026. [CrossRef]
- Harvey, J.A., Tougeron, K., Gols, R., Heinen, R., Abarca, M., Abram, P.K., Chown, S.L. Scientists’ warning on climate change and insects. Ecol Monogr. 2023; 93(1):e1553. [CrossRef]
- Godfray, H.C.J. Parasitoids: behavioral and evolutionary ecology. Princeton, NJ: Princeton University Press; 1994.
- Le Lann, C., Lodi, M., Ellers, J. Thermal change alters the outcome of behavioural interactions between antagonistic partners. Ecol Entomol. 2014; 39(5):578–588. [CrossRef]
- Seehausen, M.L., Cusson, M., Régnière, J., Bory, M., Stewart, D., Djoumad, A., Martel, V. High temperature induces downregulation of polydnavirus gene transcription in lepidopteran host and enhances accumulation of host immunity gene transcripts. J Insect Physiol. 2017; 98:126–133. [CrossRef]
- Moore, M.E., Kester, K.M., Kingsolver, J.G. Rearing temperature and parasitoid load determine host and parasitoid performance in Manduca sexta and Cotesia congregata. Ecol Entomol. 2019; 44(5):519–528. [CrossRef]
- Schreven, S.J.J., Frago, E., Stens, A., de Jong, P.W., van Loon, J.J.A. Contrasting effects of heat pulses on different trophic levels: an experiment with a herbivore–parasitoid model system. PLoS ONE. 2017; 12(4):e0176704. [CrossRef]
- Machekano, H., Mvumi, B.M., Nyamukondiwa, C. Loss of coevolved basal and plastic responses to temperature may underlie trophic level host–parasitoid interactions under global change. Biol Control. 2018; 118:44–54. [CrossRef]
- Mutamiswa, R., Chidawanyika, F., Nyamukondiwa, C. Comparative assessment of the thermal tolerance of spotted stemborer and its larval parasitoid. Insect Sci. 2018; 25(5):847–860. [CrossRef]
- Moore, M.E., Hill, C.A., Kingsolver, J.G. Differing thermal sensitivities in a host–parasitoid interaction: high, fluctuating developmental temperatures produce dead wasps and giant caterpillars. Funct Ecol. 2021; 35:675–685. [CrossRef]
- Lange, S., Volkholz, J., Geiger, T., Zhao, F., Vega, I., Veldkamp, T., et al. Projecting exposure to extreme climate impact events across six event categories and three spatial scales. Earth’s Future. 2020; 8(11):e2020EF001616. [CrossRef]
- Ranasinghe, R., Ruane, A.C., Vautard, R., Arnell, N., Coppola, E., Dessai, S., et al. Climate change information for regional impact and risk assessment. Clim Change. 2021; 1767(14):1926. DOI: 10.1017/9781009157896.014.
- Pörtner, H.O. Climate impacts on organisms, ecosystems and human societies: integrating OCLTT into a wider context. J Exp Biol. 2021; 224(Suppl_1):jeb238360. [CrossRef]
- Parmesan, C., Morecroft, M.D., Trisurat, Y., Adrian, R., Anshari, G.Z., Arneth, A., et al. Terrestrial and Freshwater Ecosystems and Their Services. In Climate Change 2022: Impacts, Adaptation and Vulnerability: Working Group II Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 197-378). Cambridge University Press; 2023. pp. 197–377. [CrossRef]
- Martin, R.A., da Silva, C.R., Moore, M.P., Diamond, S.E. When will a changing climate outpace adaptive evolution?. WIREs Clim Change. 2023; 14(6):e852. [CrossRef]
- Sgrò, C.M., Terblanche, J.S., Hoffmann, A.A. What can plasticity contribute to insect responses to climate change?. Annu Rev Entomol. 2016; 61(1):433–451. [CrossRef]
- Logan, M.L., Cox, C.L. Genetic constraints, transcriptome plasticity, and the evolutionary response to climate change. Front Genet. 2020; 11:538226. [CrossRef]
- Bonamour, S., Chevin, L.-M., Charmantier, A., Teplitsky, C. Phenotypic plasticity in response to climate change: the importance of cue variation. Phil Trans R Soc B. 2019; 374:20180178. [CrossRef]
- Le Lann, C., Van Baaren, J., Visser, B. Dealing with predictable and unpredictable temperatures in a climate change context: the case of parasitoids and their hosts. J Exp Biol. 2021; 224(Suppl_1):jeb238626. [CrossRef]
- Jerbi-Elayed, M., Foray, V., Tougeron, K., Grissa-Lebdi, K., Hance, T. Developmental temperature affects life-history traits and heat tolerance in the aphid parasitoid Aphidius colemani. Insects. 2021; 12(10):852. [CrossRef]
- Kelly, M. Adaptation to climate change through genetic accommodation and assimilation of plastic phenotypes. Phil Trans R Soc B. 2019; 374(1768):20180176. [CrossRef]
- Armarego-Marriott, T. Climatic selection and gene expression plasticity. Nat Clim Chang. 2021; 11(1):4. [CrossRef]
- Castellanos, N.L., Bueno, A.F., Haddi, K., Silveira, E.C., Rodrigues, H.S., Hirose, E., Smagghe, G., Oliveira, E.E. The fitness and economic benefits of rearing the parasitoid Telenomus podisi under fluctuating temperature regime. Neotrop Entomol. 2019; 48:934–948. [CrossRef]
- Kawecki, T.D., Ebert, D. Conceptual issues in local adaptation. Ecol Lett. 2004; 7(12):1225–1241. [CrossRef]
- Tougeron, K., Van Baaren, J., Llopis, S., Ridel, A., Doyon, J., Brodeur, J., Le Lann, C. Disentangling plasticity from local adaptation in diapause expression in parasitoid wasps from contrasting thermal environments: a reciprocal translocation experiment. Biol J Linn Soc. 2018; 124(4):756–764. [CrossRef]
- Van Emden, H.F., Harrington, R., editors. Aphids as Crop Pests. Wallingford, UK: CABI; 2017.
- Wajnberg, E., Bernstein, C., Van Alphen, J., editors. Behavioral Ecology of Insect Parasitoids: From Theoretical Approaches to Field Applications. Chichester, UK: Wiley; 2008.
- Le Ralec, A., Anselme, C., Outreman, Y., Poirié, M., van Baaren, J., Le Lann, C., van Alphen, J.J.M. Evolutionary ecology of the interactions between aphids and their parasitoids. C R Biol. 2010; 333:554–565. [CrossRef]
- Rakhshani, E., Starý, P. Aphid parasitoids: Aphidiinae (Hym., Braconidae). In: Farazmand, H., editor. Biological Control of Insect and Mite Pests in Iran. Cham: Springer; 2021. p. 333–399.
- Starý, P. The Aphidiidae of Chile (Hymenoptera, Ichneumonoidea, Aphidiidae). Dtsch Entomol Z. 1995; 42(1):113–138. [CrossRef]
- Tomanović, Z., Petrović, A., Mitrović, M., Kavallieratos, N.G., Starý, P., Rakhshani, E., Popović, A., Shukshuk, A., Ivanović, A. Molecular and morphological variability within the Aphidius colemani group with redescription of Aphidius platensis Brethes. Bull Entomol Res. 2014; 104(1):1–14. [CrossRef]
- Nieto, J.M., Fuentes-Contreras, E., Castro, M.C., Aldea, M.P., Ortego, J., Mier, P.D. Catálogo de los áfidos (Hemiptera: Aphididae) de Chile, con plantas hospedadoras y distribuciones regional y provincial. Graellsia. 2016; 72:e050. [CrossRef]
- Alvarez-Baca, J. K., Alfaro-Tapia, A., Lavandero, B., Le Lann, C., & Van Baaren, J. Suitability and profitability of a cereal aphid for the parasitoid Aphidius platensis in the context of conservation biological control of Myzus persicae in orchards. Insects, 2020. 11(6), 381. [CrossRef]
- Alfaro-Tapia A, Alvarez-Baca JK, Tougeron K et al Overwintering strategies and life-history traits of different populations of Aphidius platensis along a latitudinal gradient in Chile. Entomol. Gen. 2021. [CrossRef]
- Alfaro-Tapia, A., Alvarez-Baca, J. K., Tougeron, K., Van Baaren, J., Lavandero, B., & Le Lann, C. Composition and structure of winter aphid–parasitoid food webs along a latitudinal gradient in Chile. Oecologia 2022. 200(3), 425-440. [CrossRef]
- IPCC, Sections. In: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (eds.)]. IPCC, 2023, Geneva, Switzerland, pp. 35-115. [CrossRef]
- ODEPA. Panorama de la agricultura chilena. Oficina de Estudios y Políticas Agrarias, Santiago, Chile; 2019. Disponible en: https://www.odepa.gob.cl/wp-content/uploads/2019/09/panorama2019Final.pdf.
- Nicol, C.M.Y., Mackauer, M. The scaling of body size and mass in a host–parasitoid association: influence of host species and stage. Entomol Exp Appl. 1999; 90(1):83–92. [CrossRef]
- Dion, E., Zélé, F., Simon, J.-C., Outreman, Y. Rapid evolution of parasitoids when faced with the symbiont-mediated resistance of their hosts. J Evol Biol. 2011; 24(4):741–750. [CrossRef]
- Jerbi-Elayed, M., Tougeron, K., Grissa-Lebdi, K., Hance, T. Effect of developmental temperatures on Aphidius colemani host-foraging behavior at high temperature. J Therm Biol. 2022; 103:103140. [CrossRef]
- Piticar, A. Changes in agro-climatic indices related to temperature in Central Chile. Int J Biometeorol. 2019; 63(4):499–510. [CrossRef]
- Harvey, J.A., Cloutier, J., Visser, B., et al. The effect of different dietary sugars and honey on longevity and fecundity in two hyperparasitoid wasps. J Insect Physiol. 2012; 58:816–823. [CrossRef]
- Charles, J.J., Paine, T.D. Fitness effects of food resources on the polyphagous aphid parasitoid, Aphidius colemani Viereck. PLoS ONE. 2016; 11(1):e0147551. [CrossRef]
- Benelli, G., Giunti, G., Tena, A., Desneux, N., Caselli, A., Canale, A. The impact of adult diet on parasitoid reproductive performance. J Pest Sci. 2017; 90:807–823. [CrossRef]
- Antolin, M.F., Bjorkstein, T.A., Vaughn, T.T. Host-related fitness trade-offs in a presumed generalist parasitoid, Diaeretiella rapae (Hymenoptera: Aphidiidae). Ecol Entomol. 2006; 31:242–254. [CrossRef]
- Charnov, E.L., Los-den Hartogh, R.L., Jones, W.T., van den Assem, J. Sex ratio evolution in a variable environment. Nature. 1981; 289(5793):27–33. [CrossRef]
- Colinet, H., Boivin, G., Hance, T. Manipulation of parasitoid size using the temperature-size rule: fitness consequences. Oecologia. 2007; 152:425–433. [CrossRef]
- Zepeda-Paulo, F.A., Ortiz-Martínez, S.A., Figueroa, C.C., Lavandero, B. Adaptive evolution of a generalist parasitoid: implications for the effectiveness of biological control agents. Evol Appl. 2013; 6(6):983–999. [CrossRef]
- Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., & White, J. S. S. Generalized linear mixed models: a practical guide for ecology and evolution. Trends in ecology & evolution, 2009. 24(3), 127-135. [CrossRef]
- Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Maechler M, Bolker BM “glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling.” The R Journal, 2017. 9(2), 378–400.
- The jamovi project (2024). jamovi. (Version 2.6) [Computer Software]. Retrieved from https://www.jamovi.org.
- R Core Team (2024). R: A Language and environment for statistical computing. (Version 4.4) [Computer software]. Retrieved from https://cran.r-project.org. (R packages retrieved from CRAN snapshot 2024-08-07).
- Box, G.E.P., Cox, D.R. An analysis of transformations. J R Stat Soc B. 1964; 26:211–243. [CrossRef]
- Venables, W.N., Ripley, B.D. Modern Applied Statistics with S. 4th ed. New York: Springer; 2002.
- Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-162; 2021.
- Lenth R (2025). emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version1.10.6090003. https://rvlenth.github.io/emmeans/, https://rvlenth.gith b.io/emmeans/.
- Fox, J., Weisberg, S. An R Companion to Applied Regression. 3rd ed. Thousand Oaks, CA: Sage; 2019.
- Lüdecke, D., Ben-Shachar, M., Patil, I., Waggoner, P., Makowski, D. performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J Open Source Softw. 2021; 6(60):3139. [CrossRef]
- Cohen, A.C. Ecology of insect rearing systems: a mini-review of insect rearing papers from 1906–2017. Adv Entomol. 2018; 6(2):86. doi: 10.4236/ae.2018.62008.
- Herren, P., Hesketh, H., Meyling, N.V., Dunn, A.M. Environment–host–parasite interactions in mass-reared insects. Trends Parasitol. 2023; 39(7):588–602. [CrossRef]
- Lommen, S.T., de Jong, P.W., Pannebakker, B.A. It is time to bridge the gap between exploring and exploiting: prospects for utilizing intraspecific genetic variation to optimize arthropods for augmentative pest control—a review. Entomol Exp Appl. 2017; 162(2):108–123. [CrossRef]
- Buckley, L.B., Schoville, S.D., Williams, C.M. Shifts in the relative fitness contributions of fecundity and survival in variable and changing environments. J Exp Biol. 2021; 224(Suppl_1):jeb228031. [CrossRef]
- Le Lann, C, Wardziak, T, van Baaren, J, van Alphen, JJM. Thermal plasticity of metabolic rates linked to life-history traits and foraging behaviour in a parasitic wasp. Functional Ecology 2011. 25:641-651. [CrossRef]
- Logan, M.L., Cox, R.M., Calsbeeka, R. Natural selection on thermal performance in a novel thermal environment. PNAS 2014; 33:14165–14169. doi/10.1073/pnas.1404885111.
- Ismaeil, I., Doury, G., Desouhant, E., Dubois, F., Prevost, G. and Couty, A. (2013). Trans-generational effects of mild heat stress on the life history traits of an aphid parasitoid. PLOS One 8, 1-9. [CrossRef]
- Atkinson, D. (1994) Temperature and organism size—a biological law for ectotherms. Advances in Ecological Research, 25, 1–58. [CrossRef]
- Sinclair, B.J., Marshall, K.E., Sewell, M.A., Levesque, D.L., Willett, C.S., Slotsbo, S., et al. Can we predict ectotherm responses to climate change using thermal performance curves and body temperatures?. Ecol Lett. 2016; 19(11):1372–1385. [CrossRef]
- Giri, M.K., Pass, B.C., Yeargan, K.V., Parr, J.C. Behavior, net reproduction, longevity, and mummy-stage survival of Aphidius matricariae. Entomophaga. 1982; 27:147–153. [CrossRef]
- Zamani, A.A., Talebi, A., Fathipour, Y., Baniameri, V. Effect of temperature on life history of Aphidius colemani and Aphidius matricariae, two parasitoids of Aphis gossypii and Myzus persicae. Environ Entomol. 2007; 36(2):263–271. [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [CrossRef]
Figure 1.
Influence of temperature-rearing regimens (Constant: 20°C and fluctuating at day: 25°C; at night: 17°C) on: (A) probability density of number of mummified aphids and (B) the mean (±SE) number of mummified aphids (mummies)/female by 24hr and (C) mean (± SE) emergence rate at generation four of mass-rearing of the aphid parasitoid Aphidius platensis.
Figure 1.
Influence of temperature-rearing regimens (Constant: 20°C and fluctuating at day: 25°C; at night: 17°C) on: (A) probability density of number of mummified aphids and (B) the mean (±SE) number of mummified aphids (mummies)/female by 24hr and (C) mean (± SE) emergence rate at generation four of mass-rearing of the aphid parasitoid Aphidius platensis.
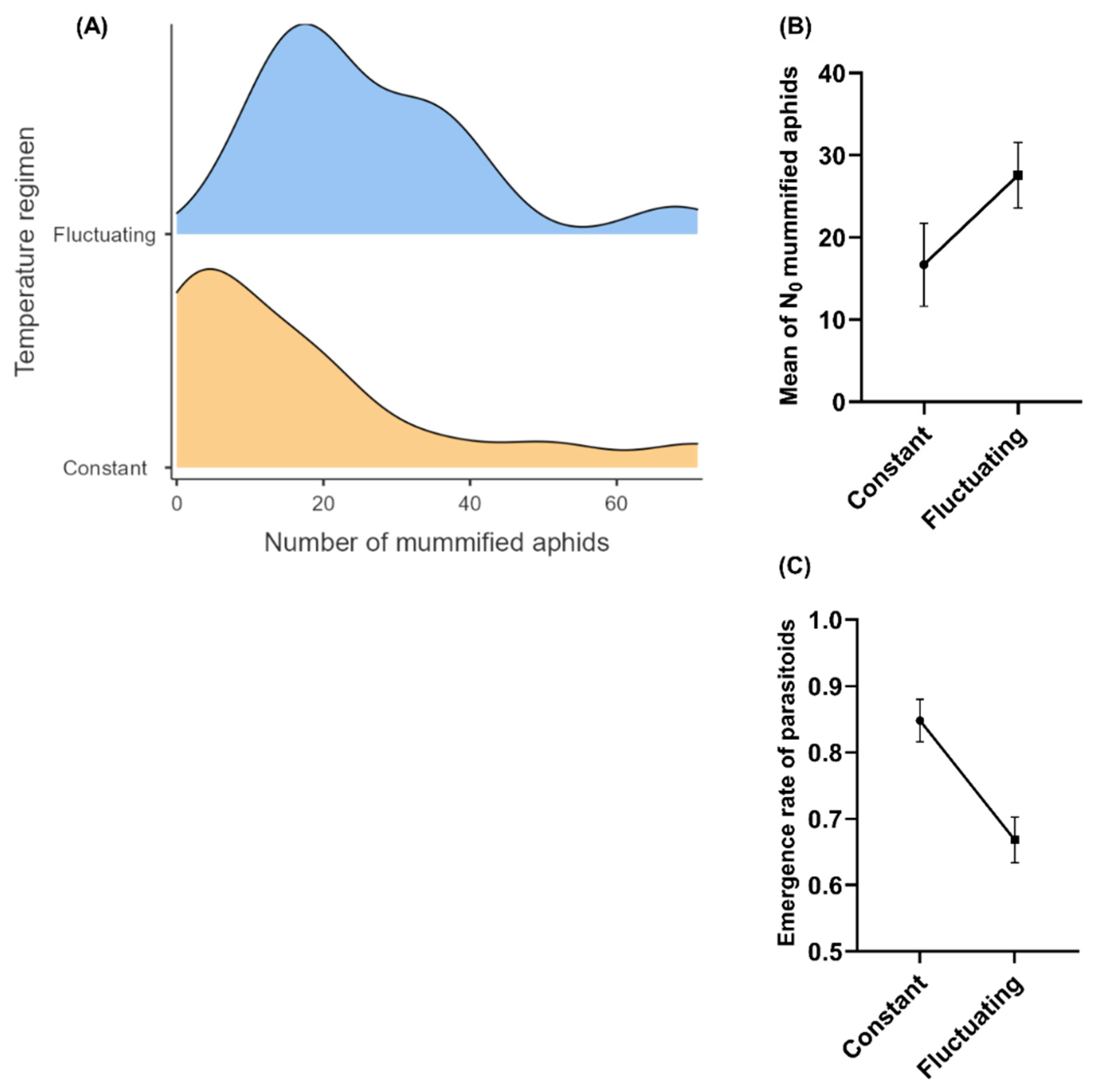
Figure 2.
Influence of rearing temperature regimens (constant: 20°C and fluctuating at day: 25°C; at night: 17°C) on (A) the mean (± SE) development time (days); (B) mean (± SE) hind-tibia length (mm) of parasitoid progeny; and (C) mean (± SE) developmental growth rate (mm day-1) at generation four of mass-rearing of the aphid parasitoid Aphidius platensis.
Figure 2.
Influence of rearing temperature regimens (constant: 20°C and fluctuating at day: 25°C; at night: 17°C) on (A) the mean (± SE) development time (days); (B) mean (± SE) hind-tibia length (mm) of parasitoid progeny; and (C) mean (± SE) developmental growth rate (mm day-1) at generation four of mass-rearing of the aphid parasitoid Aphidius platensis.
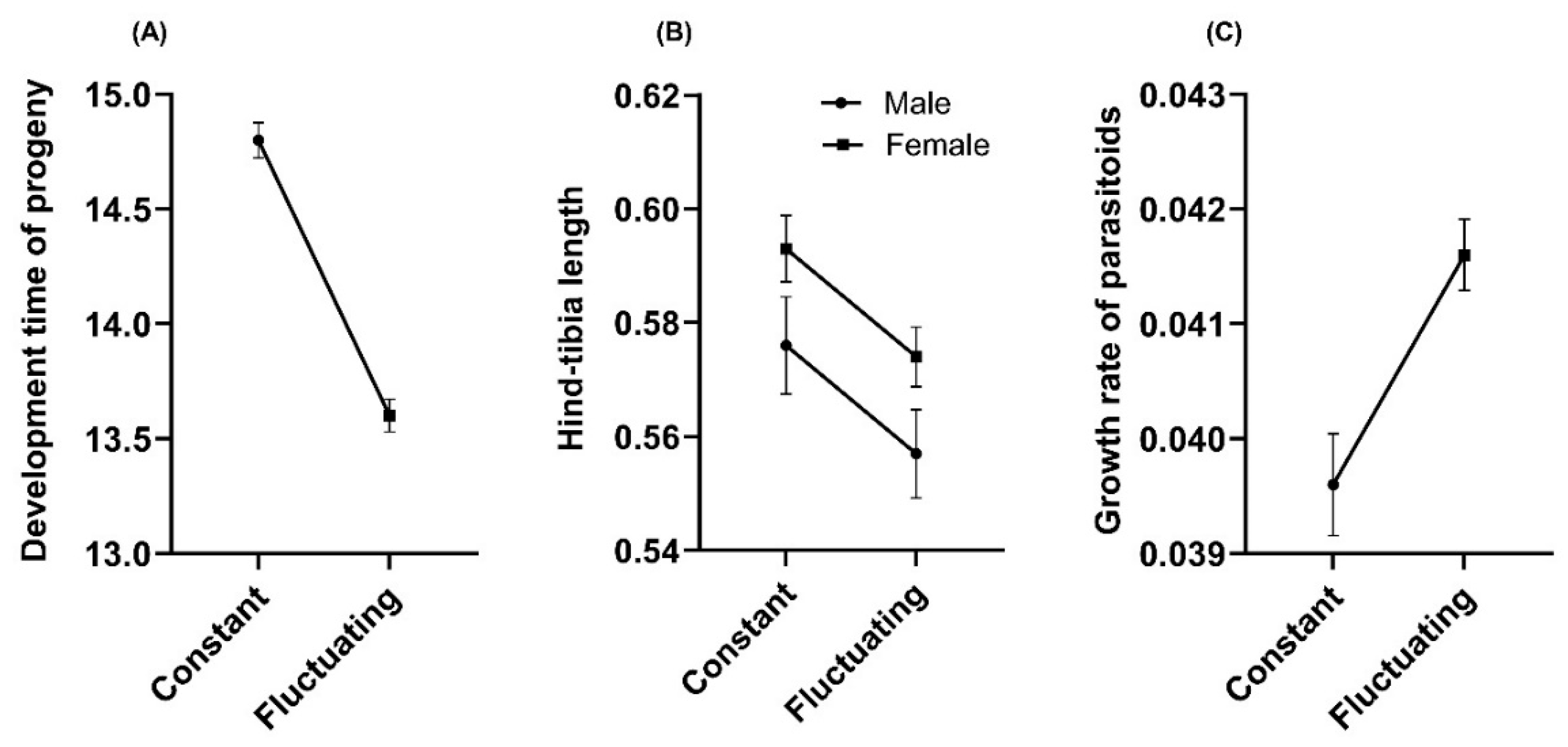
Figure 3.
Effect of rearing temperature regimens (constant: 20°C, and fluctuating at day: 25°C; at night: 17°C) on the mean (± SE) number of mummified aphids/female by 24 hr. under contrasting thermal conditions (Cold: 10°C / Standard: 20°C / Stress heat: 28°C) at generation four of mass-rearing of the aphid parasitoid Aphidius platensis.
Figure 3.
Effect of rearing temperature regimens (constant: 20°C, and fluctuating at day: 25°C; at night: 17°C) on the mean (± SE) number of mummified aphids/female by 24 hr. under contrasting thermal conditions (Cold: 10°C / Standard: 20°C / Stress heat: 28°C) at generation four of mass-rearing of the aphid parasitoid Aphidius platensis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



