
Submitted:
21 July 2025
Posted:
22 July 2025
You are already at the latest version
Abstract
In order to investigate the effect of Zataria multiflora Bioss. extract and nano-extract on morphophysiological, phytochemical indices, yield, and essential oil compositions of basil (Ocimum basilicum L.var. Genovese) under salinity stress (0, 25, 50, and 100 mM NaCl), an experiment was conducted as split-plot design in a basic block with complete randomization with three replications. The results of this study showed that the highest levels of root and shoot fresh weight, total chlorophyll, total phenol, flavonoid, antioxidant activity (DPPH and FRAP), proline, catalase enzyme and phenol alanine ammonia lyase (PAL) enzyme were observed in the Z. multiflora nanoextract treatment. The highest level of potassium was observed in conditions without salinity stress and foliar spraying with Z. multiflora extract. The highest level of O. basilicum L.var. Genovese sodium was observed in the 100 mM salinity treatment and the lowest level was recorded in the nanoextract treatment in conditions without salinity stress. In this study, the results of essential oil analysis indicated the identification of 39 compounds in which linalool, eugenol, carotenoid, methyl chavicol, A-Humulene and menthol were identified as the main compounds. In this study, the effect of nano-extract on the content of essential oil compounds was evident. The results of the study showed that there was a positive correlation between the traits of fresh and dry weight of shoots, fresh and dry weight of roots, total chlorophyll and potassium. While sodium showed a negative correlation with the traits of fresh and dry weight of shoots, fresh and dry weight of roots, total chlorophyll and potassium. Principal component analysis showed that the combination of the first two components accounted for 89.28% of the variations. Among all treatments, Z. multiflora nanoextract, while moderating the effects of stress, showed the highest efficiency in improving the morphophysiological, biochemical traits, and essential oil content and secondary metabolites of O. basilicum L.var. Genoves.

Keywords:
basil
; hydroponic
; growth
; nanoparticle
; salinity
1. Introduction
In classifying the most popular aromatic plants, basil (Ocimum basilicum L.) easily surpasses the rest [1]. Its versatile use as medicinal herb in preservatives, culinary applications, and pesticides, food flavorings, cosmetic, industrial products, and pharmaceutical has justifiably earned it the title of “king of herbs” [2,3,4]. Currently, O. basilicum is the most important commercial species of its genus and among the most popular and widespread plants worldwide [5]. Basil is an annual herbaceous plant from the Lamiaceae family, known worldwide for its aroma and fragrance [6] and it commonly consumed fresh or processed and is also used to extract essential oils [7]. Due to its genetic diversity, it differs in morphological traits and chemical and aromatic composition. Nutritionally, basil has significant health-promoting properties that have attracted the attention of consumers due to its high content of minerals, vitamins, antioxidants, and low caloric value [1]. The cultivar Genovese, which has a high content of linalool and eugenol and the absence of estragole, has an undeniable aroma and flavor that distinguishes it from other basil cultivars [8] and is widely used in medicine, perfumery, and cooking Due to its high content of secondary metabolites, especially rosmarinic and caffeic acids, it has a wide range of biological activities and extensive applications in the food industry [9]. Eugenol has anesthetic, antioxidant, and anti-inflammatory properties [10] and linalool is used in perfumery [11] and exhibits antimicrobial activity [12].
Salinity is considered a global limiting factor for agriculture, with about one-fifth of irrigated lands suffering from secondary salinity [13]. In plants, the response to salinity stress involves multidimensional changes at the molecular, cellular, and physiological levels, which usually lead to reduced yield and, in the most severe cases, growth arrest or death depending on the type, intensity, and duration of the stress [14,15,16]. Salinity stress is a concern for both irrigated and non-irrigated lands, affecting freshwater quality, soil health, biodiversity, and ultimately society [13], and climate change is predicted to exacerbate salinity problems in agriculture [5]. Therefore, it is essential to investigate and possibly adapt horticultural production systems to saltier conditions. Horticultural crops continually encounter significant abiotic stresses, leading to various effects that exacerbate global food scarcity. Thus, balancing horticultural yield with food demand necessitates the adoption of strategies that enhance plant adaptation to these challenging conditions. One of the greatest challenges facing the world today is the scarcity of water resources, which has reduced water availability for both human and agricultural use due to low rainfall levels, poor distribution, high evapotranspiration, and drought. In order to overcome water scarcity, water reuse and groundwater use are the main guarantees of water availability in the semi-arid region, especially during drought periods [17]. The increase in the world population and the need to ensure food security have doubled the importance of producing agricultural products such as fruits and vegetables using wastewater and unconventional waters (such as salt water) in order to preserve the world’s water resources [18]. Therefore, it seems that providing food for different countries under new cultivation conditions such as floating is important both in terms of freshwater shortages and salinization of the world’s water resources, and in terms of the possibility of using unconventional waters in the agricultural sector. In these circumstances, environmentally friendly solutions in mitigating salt stress for enhanced plant growth and performance seems absolutely vital.
Modern agriculture has to face many challenges, including food security, sustainable use of natural resources, and farming under harsh environmental conditions [19]. In this context, cultivation under controlled conditions has been a pivotal part of agriculture, especially for horticultural crops that are more susceptible to adverse conditions, while increasing crop yields by increasing resource use efficiency [20]. Floating is a technology in which plants are grown in a floating bed or mat on the water surface [21]. The aerial parts of the plants grow and remain above the water, where the plant roots float below the bed and are suspended in the water instead of being rooted in a solid substrate [22]. Roots absorb nutrients directly from water and form a hanging network of roots covered with biofilms, which in turn act as a filter and provide a large bioactive surface area for biochemical transformation and physical processes of pollutants [23]. Floating cultivation has attracted the attention of researchers by offering a simpler environmental technology, with lower maintenance costs and no need for additional growing media [24]. Among the different cultivation systems, floating has numerous advantages compared to soil cultivation or other soilless systems, especially for leafy vegetable production [25]. It may also improve the phytochemical-pharmacological properties of the plant by stimulating the secondary metabolism of the plant through appropriate manipulation of mineral nutrition [19].
Recently, there has been a growing interest in growing basil in soilless greenhouses, which provide the right conditions to maximize production. Because basil has high yields and good product quality when grown in a soilless system [26,27]. Rakocy et al. [28] reported that aquaponic basil produced a higher yield (1.8 kg/m2) than field basil (0.6 kg/m2). Basil, when grown in a soilless system, offers high yields and good crop quality [27]. Basil is a salinity-sensitive plant [7].
Nowadays, the use of synthetic antioxidants compounds in medicine (Acetylcarnitine, Allopurinol, Butylated hydroxyanisole and etc), agriculture, and pharmaceutical industries has become very popular, but numerous studies indicate the toxicity of these antioxidants [29]. For this reason, finding natural metabolites, especially from plant sources, is important [30]. On the other hand, it has been shown that antioxidants of natural origin are more stable [31]. Despite their production in small quantities by plants, secondary metabolites improve the organism’s potential to resist biotic and abiotic stress conditions, including salinity, temperature fluctuations, osmotic and drought stresses, metal toxicity, and pathogen attack [32]. Referring to the role of secondary metabolites as plant antioxidants, it can be stated that natural antioxidants have a major role in neutralizing excess free radicals (ROS) [33]. This system includes enzymatic and non-enzymatic types [34]. Considering that plants are one of the important sources of antioxidants, species rich in antioxidant compounds can protect cells from oxidative damage (including salt stress and the resulting oxidative damage) [35]. These antioxidants are polyphenolic compounds found in different parts of plants [36]. Recent findings have shown that plant antioxidants, especially pure extracts containing compounds such as polyphenols, flavonoids, amino acids, carotenoids, and cytokinins, play a key role in improving plant growth under stress conditions by modulating endogenous phytohormone levels and increasing carbohydrate accumulation [37]. A recent study in a plant of the Lamiaceae family (Thymbra spicata L.) showed that compounds such as polyphenols along with carvacrol, gamma-terpinene, para-cymene, caryophyllene, myrcene, and alphaterpinene play an important role in modulating the negative effects of salinity [38,39].
Researchers have found that different species of Lamiaceae family are rich in antioxidants [40,41,42]. The medicinal uses of Lamiaceae family are mostly related to the presence of two classes of secondary metabolites including phenolic compounds and monoterpenoids. Phenolics are the largest group of secondary metabolites, with phenols having a significant content in mint species, showing specific biological activities such as antioxidant effects [36,43]. Within the same family, the extract of Shirazi thyme (Zataria multiflora Bioss.) has a high content of phenols and flavonoids [44].
Z. multiflora is native in Southwest Asia (Iran, Afghanistan, and Pakistan) that contains high levels of carvacrol and thymol [45]. The essential oil of Z. multiflora is rich in bioactive compounds that show specific biological effects (anti-inflammatory, antipyretic, antioxidant, allelopathy, antiulcer, antimicrobial and hepatoprotective) [45,46] and contains high contents of carvacrol (4.99 - 46.22%) and thymol (14.8 - 70.12%) [45]. In this regrads, a study designed to investigate the effects of Thymbra spicata extract at concentrations of 0% (C), 10% (TS1), and 20% (TS2), along with mycorrhizal fungus biofertilizer at a rate of 50 g/2.5 kg of soil, were evaluated on Rosmarinus officinalis plant under salinity stress conditions. The results showed as salinity stress increased, the yield and growth characteristics of the plants declined. However, the applied treatments effectively mitigated the negative effects of salinity. Among all the treatments, the combined application of TS2 and MF proved to be the most effective in enhancing the morphophysiological and biochemical characteristics of rosemary plants. This treatment not only boosted the production of essential oils and secondary metabolites but also mitigated the detrimental effects of salinity stress [39].
Recent research has shown the promising potential of nanotechnology to improve the agricultural sector by increasing the efficiency of agricultural inputs and providing solutions to agricultural and environmental problems to improve productivity and food security [47,48]. Promising results have been reported in reducing the effects of stress after the application of nanoparticles in various plants [49,50]. Nanoparticles are atomic or molecular assemblies with dimensions between 1 and 100 nm that have different physicochemical properties compared to their bulk materials. Their uptake and transport vary depending on the plant species, size, chemical composition, structure, and strength of the nanoparticles [51]. Converting materials to the nanoscale leads to changes in their physicochemical, biological, and catalytic properties and contributes to better solubility and greater penetration of cell membranes [52,53]. In a study, the biosynthesis of Ag-nano particles from extracts of Marrubium alysson and Torilis arvensis investigated on wheat plants under drought stress, the results showed that natural and nano extracts significantly increased CAT and GPX as well as soluble proteins. Phenolics, flavonoids, soluble sugars, H2O2 and MDA content were all significantly reduced. This study believed that priming wheat with natural and nano extracts improved drought tolerance through the utilization of its metabolites, which included soluble sugars, phenolics and flavonoids, and accumulated in other metabolites such as lignin, starch and flavonan to enhance plant tolerance and reduce oxidative damage. Furthermore, the results showed that nano extracts of Torilis arvensis and Marrubium alysson were more effective than plant extracts because they were separated from each other in PCA analysis [54].
In recent decades, professional basil cultivation has gradually shifted towards soilless systems in controlled and closed environments [27]. Global warming, drought, and changes in rainfall patterns and water consumption have led to a sharp decline in water resources. Given the climate situation, salinity should be used as an opportunity instead of using fresh water. As a result, one of the solutions proposed to manage these conditions is the use of unconventional waters (including saltwater, wastewater, and sewage), especially in the agricultural sector. Therefore, finding crop management techniques that increase resistance, plant water consumption efficiency, and consequently plant growth under abiotic stress conditions, especially salinity stress, will be very useful. Therefore, in this study, the effect of Shirazi thyme (Z. multiflora) extract and nano-extract on salt stress tolerance of O. basilicum ‘Genovese’ under floating system was investigated.
2. Materials and Methods
2.1. Plant Material
First, to obtain identical plants, O. basilicum ‘Genovese’ seeds were obtained from Pakan Seed Company, Iran. Then, the seeds were sterilized with 5% sodium hypochlorite for 30 seconds, and then washed several times with distilled water, and after drying, they were sown in seedling trays. When the seedlings reached the four-leaf stage (14 days later), they were transferred to plastic trays containing water and nutrients and placed in suitable conditions (with polyethylene cover and average night temperature of 15-20°C and day temperature of 20-25°C and relative humidity of 60-70%) for ten days to establish.
2.2. Experiment Design
This experiment was conducted in a split-plot design as a basic block with complete randomization with three replications (each replication had five plants) in the Medicinal Plants Research Greenhouse, Department of Horticultural Sciences, Faculty of Agriculture, Ilam University at longitude 46˚22’E, latitude 33˚39’N and altitude 1446 m above sea level. The experimental treatments included salinity stress treatment at four levels (including 0 (S1), 25 (S2), 50 (S3), and 100 mM NaCl (S4)) in the form of water solution along with irrigation and treatment of foliar spray with Z. multiflora extract at three levels (including control (E1), Z. multiflora ethanol extract (E2), and Z. multiflora nano ethanol extract (E3)) in the form of a foliar spray, which were applied three times during the experiment (10, 20 and 30 days after transplant). The plants were fed using a modified Hogland nutrient solution (N (136), P (31), K (215), Ca (84), Mg (24), Fe (0.9), B (0.4), Mn (0.14), Zn (0.32), Cu (0.05), and Mo (0.05) mg L-1) as an irrigation fertilizer and harvested after 60 days. For floating basil cultivation, each plastic tray was filled with 50 liters of nutrient solution and an air pump (output= 280 L/min) with 16 mm drip irrigation pipe, dropper 2 liters per hour, air stone, and spaghetti tubes and required fitting were used to provide oxygen to the nutrient solution.
2.3. The Extraction Methods
To prepare the ethanol extract of Z. multiflora, branches in the flowering stage were dried, powdered, and then extracted (Plants were collected from the research farm of Ilam University) and 150 g of each sample was soaked in one liter of distiled water and 50% ethanol (v/v). The resulting solutions were placed on a shaker for 48 h and then filtered using Whatman filter paper number 1. Then, to separate the ethanol, the solutions were placed in the open air for 72 h and the resulting extract was used as a stock solution for spraying and preparing nanoextracts. The method of Ghazy et al. [55] was used to prepare the nanoextract. For this purpose, Z. multiflora extract was emulsified in the aqueous phase using an ultrasonic homogenizer (Q700 sonicator, Qsonica, USA). Specifically, 5 mL of the bulk extract was added through a 0.45 μm syringe filter to 10 mL of H2O containing 5% Tween 80 (based on the weight of the extract). Sonication was performed using a ½-inch tip for 3 min at 50% amplitude, under ice cooling. To prevent contamination, the nanoemulsion was covered with perforated aluminum foil and stirred at room temperature for 24 h to remove ethanol. Finally, the nanoemulsion was stored in glass vials at room temperature in the dark. Figure 1 shows the TEM image of Z. multiflora nano extract.
2.4. Measurement and Sampling
In June 2024 (90 days after planting) measurement and sampling (freeze-dried) were conducted to perform physiological, biochemical, and secondary metabolite tests.
2.5. Vegetative Traits
The plants were harvested from the crown and divided into smaller pieces with garden scissors, and the fresh weight of the roots and shoots was immediately measured with a scale with an accuracy of 0.01 g. The dry weight of the shoots and roots was also measured after drying in the shade for one week.
2.6. Total Chlorophyll Content
To determine the chlorophyll content, the Arnon method [56] was used with slight modifications. 0.5 g of leaf sample was weighed and placed in a mortar. Then, five milliliters of 80% acetone solution was added to the mortar and the plant sample was crushed well. Next, the crushed leaf mixture and acetone were transferred to a Falcon tube. The tube containing the leaf sample was centrifuged for five minutes at 5,000 rpm (Prismr, Labnet, USA). Then, the solution was transferred to a 100 ml volumetric flask using a funnel and filter paper and made up to volume. Using a spectrophotometer (Specord 50, Analytic Jena AG, Germany), the light absorbance of the solution was read at wavelengths of 663 and 645 nm and finally, the total chlorophyll content was calculated in mg/g of fresh weight of the sample using the following formula.
Total chlorophyll = [(19.3 * A663 - 0.86 * A645) V/100W] + [(19.3 * A645 - 3.6 * A663) V/100W]
2.7. Total Phenol and Flavonoid Content
Methanolic extract was used to measure antioxidant compounds. To prepare the methanolic extract, 500 mg of leaf tissue was added to 10 mL of 80% methanol as solvent and the resulting mixture was homogenized for 24 h using a shaker. Then, to separate the pure extract, the mixture was centrifuged for 10 min. The supernatant extract was stored in dark, air- and light-tight glass bottles in a refrigerator at 4°C. To measure phenol, 0.5 ml of methanolic extract was mixed with 5 ml of Folin-Ciocalteau (Merck). Then, 1 ml of 1 M sodium carbonate was added. For the blank, distilled water was used instead of the extract, and then Folin-Ciocalteau and sodium carbonate were added. This solution was used to zero the spectrophotometer (Specord 50, Analytic Jena AG, Germany). The above solution was kept in the dark for 15 minutes and the absorbance of the samples was read at a wavelength of 760 nm [57]. Flavonoids were measured with a spectrophotometer using the method of Krizek et al. [58]. Total phenol content was calculated in mg of gallic acid equivalent per g of fresh weight (mg GA/g FW) and flavonoids in mg of quercetin per g of fresh weight (mg QE/g FW) of the sample.
2.8. Proline Content
Proline was extracted from the youngest fully developed leaves using the Bates method [59]. 0.1 g of leaf tissue was ground in 10 ml of 3% sulfosalicylic acid and the resulting homogenate was centrifuged at 10,000 rpm at 4°C for 10 min. Then, in a separate tube, 2 ml of ninhydrin reagent and 2 ml of pure glacial acetic acid were added to 2 ml of the resulting extract. The tubes were then placed in a water bath at 100°C for 1 hour, and after removing them from the water bath and adding 4 ml of toluene to each tube, they were vortexed for 20 seconds. After the formation of two separate phases, the upper-colored phase was carefully separated and measured in a spectrophotometer at a wavelength of 520 nm. The proline standards dissolved in the toluene phase were poured into the cuvette of the spectrophotometer in the required amount, and the amount of proline was read at a wavelength of 520 nm and a standard curve was drawn. Then, the absorbance in the plant samples was read and the amount of proline was obtained by inserting it into the linear equation.
2.9. Catalase (CAT) and Phenylalanine Ammonia-Lyase (PAL) Enzyme Activity
First, protein extract was prepared. For this purpose, one gram of fresh tissue was ground well in a porcelain mortar containing 5 ml of 0.05 M Tris-HCl buffer with pH = 7.5. Then, it was centrifuged for 20 minutes at 12,000 rpm and 4°C. At the end of the centrifugation step, the supernatant solution (containing protein extract) was used to examine catalase activity. After the preparation of the protein extract, CAT enzyme activity was measured using the Chance and Maehly method [60] with some modifications. For this purpose, 2.5 ml of 50 mM Tris buffer pH=7 was mixed with 30 μl of hydrogen peroxidase (H2O2) and 60 μl of enzyme extract in an ice bath. The absorbance curve at 240 nm was read for one minute using a spectrophotometer. The enzyme activity was calculated as the amount of hydrogen peroxide deactivated per minute per gram of fresh leaf tissue. Phenylalanine Ammonialyase enzyme activity was measured using the Wang et al. [61] method and based on the concentration and amount of cinnamic acid produced.
2.10. Antioxidant Activity by DPPH and FRAP Methods
For measurement of DPPH radical scavenging activity a 4% DPPH radical stable solution was prepared and 2800 μL of this solution was diluted with 200 μL of the extract (final volume 3 mL) and combined in dark test tubes. The absorbance of the samples was read with a spectrophotometer at a wavelength of 517 nm [62], and the antioxidant activity by the FRAP method [63]. This method contains TPPZ (Tripyridyl-S-Triazine), FeCl3 and acetate buffer. Antioxidants that can reduce Fe3+ to Fe2+ convert the colorless TPTZ-Fe3+ complex into the TPTZ-Fe2+ complex, which is blue and its intensity can be measured at a wavelength of 593 nm. For this purpose, 250 μg/ml of the plant extract was added to a final volume of 2 ml of FRAP solution containing 10 mM TPTZ in HCl (40 mM), 20 mM ferric chloride, and 300 mM buffer with a pH of 3.6. The above sample was placed at 37°C for 10 min. The resulting color intensity was read at a wavelength of 593 nm against a blank.
2.11. Sodium and Potassium Content
The dry ash method was used to measure the sodium and potassium content. Basil leaves were dried at 80°C and then powdered. Then, 100 mg of each sample was separately burned at 560°C for 5 hours and the ash was digested in 10 mL of 1 M hydrochloric acid at 25°C for 24 hours. These samples were used to measure the sodium and potassium content by atomic absorption spectrophotometry (Shimadzu Model: AA-7000, Kyoto, Japan) [64].
2.12. Identification of Essential Oil Compounds with GC-MS
To obtain essential oil, 50 g of dry branches were mixed with 500 ml of water in a 1-liter flask and the essential oil was extracted by water distillation using a Clevenger apparatus. Then, the sample was prepared for identification of compounds preparation of GC and GC/MS spectra of the essential oil, and identification of its constituent components. Identification and evaluation of volatile compounds in basil essential oil was carried out using a gas chromatography device (GC, GMI, SCION SQ W.436, model SSL-T21, UK) connected to a SCION-MS mass spectrometer, a single quadruple spectrometer (DB-5ms, 15 m long × 0.25 mm, stationary phase layer thickness 0.25 microns), and an electron ionization source. Helium gas with a purity of 99.99% and a flow rate of 1 (min/ml) was used as the carrier gas with an ionization voltage of (eV70). The injector and interface temperatures were 280 °C and 200 °C, respectively. The mass range was between 40 and 600 (amu). The oven temperature program was the same as mentioned above for GC. Identification of compounds was done by comparing their mass spectra with the mass spectra of the internal reference library (NIST11). For injection into the device, the essential oil was first diluted with dichloromethane (0.5% solution) and then one microliter of it was injected into the device. Finally, most of the compounds were identified using the Quartz index (KI), retention time (RT), and mass spectrum [65,66].
2.14. Data Analysis and Statistical Calculations
Statistical analysis of the measured trait data was performed using SAS 9.1 software (SAS Institute, Cary, NC, USA). EXCEL software (Microsoft Office Professional Plus- 2013) was also used to draw graphs. R studio v1.3.959 was used for principal component analysis (PCA) and heat map correlation (HMC) to identify relationships between traits and treatment dispersion.
3. Results
3.1. Vegetative Traits
Based on the results, the interaction effect of salinity and foliar application of Z. multiflora extract on fresh and dry weight of shoots and roots and potassium content of O. basilicum L. var. Genovese leaves was significant at the 1% probability level and on chlorophyll and sodium content at the 5% probability level (Table 1). The highest shoot fresh weight of O. basilicum L. var. Genovess was observed in the plants treated with Z. multiflora nano extract-without salinity stress with a rate of 71.64 g. While, the lowest value was observed in the 100 mM salinity treatment-without extract stress with a rate of 12.10 g. In treatment 0 mM of NaCl salinity, nano extract increased the shoot fresh weight by 34.28% and regular extract by 8.35% compared to the control. In all salinity concentrations applied, nano extract increased shoot fresh weight more than regular extract and regular extract increased shoot fresh weight more than treatment without extract at the same salinity concentration (Table 1).
The highest shoot dry weight of O. basilicum L.var. Genoves plant was observed in the control treatment (without salinity stress) - Z. multiflora nano extract with a rate of 10.01 g. The lowest level of this index was observed in the 100 mM salinity treatment and without extract with a rate of 1.46 g. In the treatment without salinity stress, the regular extract increased the shoot dry weight by 9.53% and the nano extract by 34.36% compared to the control. In all salinity concentrations applied, the nano extract increased the shoot dry weight more than the regular extract and the treatment without extract at the same salinity concentration (Table 1).
The highest root fresh weight of O. basilicum L.var. Genoves was observed in the control treatment (without salinity stress) - Z. multiflora nano extract with a level of 103.13 g. The lowest level of this index was in the 100 mM salinity treatment and without extract with a level of 26.86 g. In the treatment without salinity stress, nano extract increased the fresh weight of roots by 34.21% and regular extract by 16.05% compared to the control. In all salinity concentrations applied, nano extract increased the fresh weight of roots compared to regular extract and regular extract increased the fresh weight of roots compared to the treatment without extract at the same salinity concentration (Table 1).
The highest dry weight of O. basilicum L.var. Genoves roots were observed in the control treatment (without salinity stress) - Z. multiflora nano extract with a level of 9.99 g. The lowest levels of this index were observed in the 100 mM salinity treatment - without extract and 100 mM - regular extract with a level of 2.80 and 3.30 g, respectively, which were not significantly different from each other. In the non-saline stress treatment, nano extract increased root dry weight by 34.63% and regular extract by 16.44% compared to the control. In all salinity concentrations applied, nano extract was observed to increase root dry weight compared to regular extract and regular extract was observed to increase root dry weight compared to the treatment without extract at the same salinity concentration (Table 1).
3.2. Total Chlorophyll
The highest total chlorophyll of O. basilicum L. var. Genovese were observed in the control treatment (without salinity stress) - Z. multiflora nano extract with a level of 8.25 mg/g FW. The lowest level of this index was in the 100 mM salinity treatment - without extract and 50 mM - without extract with a level of 3.15 and 3.87 mg g-1 FW, respectively, which were not significantly different from each other. In the non-saline stress treatment, nano extract increased total chlorophyll by 45.24% and regular extract by 30.63% compared to the control. In all salinity concentrations applied, nano extract was observed to have more total chlorophyll than regular extract, and regular extract was observed to have more total chlorophyll than the treatment without extract at the same salinity concentration (Figure 2-A).
3.3. Sodium (Na) and Potassium (K) Content
The highest Na content of O. basilicum L. var. Genovese were observed in the 100 mM salinity treatment without Z. multiflora extract with a level of 0.46 mg g-1. The lowest level of Na was obserdev in the nano extract-without salinity treatment with a level of 0.20 mg g-1. In the treatment without salinity stress, nano extract decreased Na content by 17% and regular extract by 5% compared to the control. In all salinity concentrations applied, nano extract showed lower sodium accumulation than regular extract and regular extract compared to the treatment without extract at the same salinity concentration (Figure 3-B).
The highest K content of O. basilicum L.var. Genoves were observed in the non-salinity treatment of Z. multiflora extract with a level of 6.11 mg g-1. The lowest levels of K were observed in the 100 mM salinity treatment without extract, nano-extract, and regular extract with levels of 3.59, 3.61, and 3.64 mg g-1, respectively, which were not significantly different from each other. In the non-salinity stress treatment, nano-extract increased the K content by 22.93% and regular extract by 9.05% compared to the control. In salinity concentrations applied, regular extract and nano extract showed a higher potassium accumulation rate compared to the treatment without extract at the same salinity concentration (Table 1).
3.4. Total Phenols and Flavonoid Content
Based on the results, the interaction effect of salinity and foliar application of Z. multiflora extract on the phenol and flavonoid content of O. basilicum L. var. Genovese leaves was significant at the 1% probability level (Table 2). The highest total phenols of O. basilicum L.var. Genoves were observed in the 100 mM salinity treatment in both nano-extract and regular extract of Z. multiflora, followed by the 50 mM salinity treatment - nano-extract, with 12.33, 11.17, and 10.01 mg of gallic acid per gram of fresh weight, respectively. The lowest content of phenol were observed in the control treatment (without extract and salinity level) and regular extract without salinity level, with 5.26 and 5.50 mg GA g-1 FW, respectively, which were not significantly different from each other. In the non-saline stress treatment, nano-extract increased the total phenol content by 36.12% and regular extract by 4.56% compared to the control. In all salinity concentrations applied, nano-extract was observed to have a higher total phenol content than regular extract and regular extract was observed to have a higher total phenol content than the treatment without extract at the same salinity concentration (Figure 2-A).
The highest amount of O. basilicum L. var. Genovese flavonoid in the 100 mM salinity condition were observed in the nano extract of Z. multiflora with a content of 4.44 mg QE g-1 FW. The lowest amount of flavonoid was in the control treatment (without extract and salinity level) and then the regular extract-without salinity level with content of 0.90 and 1.23 mg QE g-1 FW, respectively, which were not significantly different from each other. In the treatment without salinity stress, the nano extract increased the flavonoid content by 135.55% and the regular extract by 36.66% compared to the control. In all salinity concentrations applied, the nano extract had higher flavonoid content than the regular extract and the regular extract had higher flavonoid content than the treatment without extract at the same salinity concentration (Figure 2-B).
3.5. Proline
Based on the results, the interaction effect of salinity and foliar application of Z. multiflora extract on proline content of O. basilicum L. var. Genovese leaves was significant at the 1% probability level (Table 2). The highest proline content of O. basilicum L.var. Genoves were observed in the 100 mM salinity treatment in Z. multiflora nano extract with a level of 0.92 mg g-1 FW followed by the 50 mM salinity treatment - nano extract with a level of 0.91 mg g-1 FW which did not differ significantly from each other. The lowest level of proline was observed in the control treatment (without extract and salinity) with a level of 0.24 mg g-1 FW. In the non-saline stress treatment, nano extract increased the proline content by 125% and regular extract by 79.16% compared to the control. In all salinity concentrations applied, nano extract had a higher proline content than regular extract and regular extract than treatment without extract at the same salinity concentration (Figure 2-C).
3.6. Catalase Enzyme and Phenolalanine Ammonialyase (PAL) Enzyme
Based on the results, the interaction effect of salinity and foliar application of Z. multiflora extract on the catalase and PAL enzymes of O. basilicum L.var. Genoves was significant at the 1% and 5% probability level, respectively (Table 2).
The highest level of O. basilicum L. var. Genovese catalase enzyme was observed in 100 mM salinity treatment in Z. multiflora nano extract with a level of 0.97 μM min-1g-1FW. The lowest level of catalase was in the control treatment (without extract and salinity level) with a level of 0.22 μM min-1g-1FW. In the treatment without salinity stress, nano extract increased the level of catalase enzyme by 36.36% and regular extract by 18.18% compared to the control. In all salinity concentrations applied, nano extract was observed to have a higher level of catalase enzyme than regular extract and regular extract was observed to have a higher level of catalase enzyme than the treatment without extract at the same salinity concentration (Figure 3-A).
The highest level of O. basilicum L. var. Genovese PAL enzyme was observed in the 100 mM salinity treatment in the regular extract and nano-extract of Z. multiflora, respectively, with a level of 0.36 μgCin min-1g-1FW and 0.33. The lowest level of this index was observed in the control treatment (without extract and salinity level), extract and nano-extract without salinity levels with a level of 0.12, 0.13, and 0.14 μgCin min-1g-1FW, respectively, which were not significantly different from each other. In the treatment without salinity stress, the nano-extract increased the level of PAL enzyme by 16.66% and the regular extract by 8.33% compared to the control. In all salinity concentrations applied, the regular extract increased the level of PAL enzyme by more than the treatment without extract at the same salinity concentration. PAL enzyme content in nano-extract was higher than the regular extract in all treatments except for the 100 mM concentration, which showed a lower level than the regular extract (without a significant difference) (Figure 3-B).
3.7. Antioxidant Activity (DPPH and FARP)
Based on the results, the interaction effect of salinity and foliar application of Z. multiflora extract on the DPPH and FRAP of O. basilicum L.var. Genoves was significant at the 5% probability level (Table 2). The highest antioxidant activity of O. basilicum L.var. Genoves were observed in the 100 mM salinity treatment in both nano-extract and regular extract of Z. multiflora, followed by the 50 mM salinity treatment - nano-extract with 31.86, 30.60, and 28.21%, respectively. The lowest levels of this index were observed in the control treatment (without extract and salinity level), the treatment without extract - 25 mM salinity with 11.96 and 13.35%, respectively, which were not significantly different from each other. In the non-saline stress treatment, nano-extract increased the DPPH activity by 105.01% and regular extract by 96.07% compared to the control. In all salinity concentrations applied, nano-extract had higher antioxidant activity than regular extract and regular extract had higher antioxidant activity than the treatment without extract at the same salinity concentration (Figure 4-A).
The highest antioxidant activity (FARP) of O. basilicum L.var. Genoves were observed in the 100 mM salinity treatment and nano-extract of Z. multiflora with a rate of 0.39%, followed by 0.35% in the 50 mM salinity treatment - nano-extract, which did not differ significantly from each other. The lowest level of this index was in the control treatment (without extract and salinity level) with a rate of 0.17%. In the treatment without salinity stress, nano-extract increased the antioxidant activity by 70.58% and regular extract by 35.29% compared to the control. In all salinity concentrations applied, nano-extract had a higher antioxidant activity than regular extract and regular extract had a higher antioxidant activity than the treatment without extract at the same salinity concentration (Figure 4-B).
3.8. Identification of Essential Oil Compounds with GC-MS
In this study, the results of essential oil analysis indicated the identification of 39 compounds, in which Linalool, Eugenol, Carvone, Methyl Chavicol, A-Humulene, and Menthol were identified as the main compounds (Table 3).
The present study shows that the composition of linalool in the absence of salinity and without extract (12.3%) increased to 13.25% and 15.3% with regular extract and nanoextract, respectively. With increasing salinity to 25 mM, the percentage of linalool in the absence of extract reached 22.64%, but with the addition of regular extract and nanoextract, it showed a relative decrease (18.26% and 21.26%). At 50 mM salinity, linalool had high values at all extract levels (22.25% to 23.13%), but reached its maximum at 100 mM salinity with nanoextract (23.91%) (Table 3).
Eugenol also had a similar trend, so that with increasing salinity from zero to 50 mM, its percentage increased at all extract levels (from 2.4% at zero salinity and without extract to 7.96% at 50 mM salinity with regular extract). But at 100 mM salinity, a significant decrease was observed (4.15% with nanoextract) (Table 3).
Carvone was 15.9% in the absence of salinity and without extract, and changed little with the addition of extract. With increasing salinity to 25 mM, this compound reached 18.23% in the absence of extract, but it decreased sharply with regular extract and nanoextract (9.24% and 12.4%). This downward trend continued at 50 and 100 mM salinity, and the lowest value (7.37%) was recorded at 100 mM salinity with regular extract (Table 3).
Alpha-humulene showed the opposite behavior, so that its percentage decreased with increasing salinity and the presence of extract. For example, at zero salinity and without extract, it was 10.2%, but at 50 mM salinity with regular extract it reached 1.36%. At 100 mM salinity, this compound improved slightly (3.26% with nanoextract) (Table 3).
In contrast, menthol had an upward trend at all salinity and extract levels. It was 4.2% at zero salinity and without extract, but with increasing salinity to 100 mM and adding nanoextract, it reached 13.62%, indicating the positive effect of salinity stress and spraying of Z. multiflora extract on the accumulation of this compound (Table 3).
In this study, both alpha-humulene and methyl chavicol decreased at 25-50 mM salinity, but methyl chavicol improved at 100 mM salinity with nanoextract, while alpha-humulene showed less improvement. Linalool and menthol increased in salinity, while methyl chavicol decreased. This indicates that the plant may divert resources to other compounds. Also, eugenol peaked at 50 mM salinity (7.96%), but methyl chavicol had the minimum amount (1.05%) under the same conditions. This may indicate competitive metabolic pathways (Table 3).
In the no-salinity level (control), foliar application with nano-extract had higher methyl chavicol levels than regular extract, and regular extract had higher methyl chavicol levels than the no-salinity treatment. With increasing salinity, the methyl chavicol levels decreased significantly (Figure 5). At salinity levels of 25 and 50 mM NaCl salinity, methyl chavicol levels in the no-foliar application treatment were significantly higher than those in the regular extract and nano-extract treatments (although no significant difference was observed between the control and nano-extract treatments at the 50 mM level), but with increasing salinity levels to 100 mM salinity, the use of Z. multiflora nano-extract had more satisfactory results in both regular extract and nano-extract levels compared to the control. So that, its concentration in the nano-extract, regular extract, and control treatments in the 100 mM salinity treatment was 5.47%, 4.29%, and 2.38%, respectively, indicating the effective role of this extract in modulating severe stresses by increasing the production of methyl chavicol in O. basilicum L.var. Genoves plants (Figure 5).
Overall, the results show that salinity stress inhibits methyl chavicol synthesis, especially at 25-50 mM levels, while the application of nanoextract can compensate for the negative effects of severe salinity (100 mM) and partially restore methyl chavicol synthesis (Figure 5).
3.9. Correlation and Principal Component Analysis (PCA)
The results of the study showed that there is a positive correlation between the traits of shoot fresh and dry weight, root fresh and dry weight, total chlorophyll, and potassium, while they showed a negative correlation with the others. According to the results of this study, there is a positive correlation between total phenol, flavonoid, proline, catalase and phenylalanine ammonia-lyase enzymes, antioxidant activity, and sodium. Also, sodium showed a negative correlation with the traits of shoot fresh and dry weight, root fresh and dry weight, total chlorophyll, and potassium (Figure 6-A).
Figure 4-B, principal component analysis showed that the combination of the first two components explained 89.28% of the variations, which accordingly included 69.45% of PC1 and 24.83% of PC2. Based on this analysis, shoot and root dry weight, total chlorophyll, total phenol, flavonoids, catalase and phenylalanine ammonia-lyase enzymes, sodium, and potassium, which were most affected by salinity treatment, played a role in loading PC1. Antioxidant activity and proline, whose highest content was recorded in the Z. multiflora extract and nanoextract treatment, played a role in loading PC2 (Figure 6-B).
4. Discussion
In the present study, a decrease in the fresh and dry weight of O. basilicum L.var. Genoves shoots and roots under salinity stress were observed, which is due to osmotic effects, a decrease in osmotic potential, a decrease in water absorption, and prevention of water entry into the plant [67]. Also, by reducing turgor pressure, stomata close, photosynthesis and cell divisions decrease [68]. On the other hand, salinity increases the energy required to maintain normal cell conditions, and less energy remains to support plant growth [69]. One of the important and influential factors in the photosynthetic capacity of plants is a decrease in chlorophyll concentration, and salinity stress causes a decrease in chlorophyll, photosynthesis, and intensification of stress-related damage. Therefore, the decrease in the growth of morphological traits can be attributed to the lack of photosynthetic materials required for growth [70]. Salinity stress increases the concentration of growth regulators such as abscisic acid and ethylene, which stimulate the activity of the chlorophyllase enzyme, and thus macromolecules and chlorophyll are decomposed under the influence of this enzyme [71]. One of the strategies for dealing with the effects of stress is the use of treatments that improve plant resistance to stress, including foliar spraying with fertilizer, plant extract, etc., which in this study focused on foliar spraying with Z. multiflora extract to reduce the use of chemical fertilizers and implement sustainable agriculture, while using the capacity of medicinal plants to deal with the effects of inevitable stresses, including salinity. As mentioned in the results section, at all salinity levels, Z. multiflora extract improved the growth conditions of O. basilicum L.var. Genoves plants compared to the control at the same level. The extract causes the plant growth cycle through various mechanisms, including stimulating the activity of hormones and enzymes. Sometimes the components of the extracts contain elements such as nitrogen, phosphorus, magnesium, and other macro and micromolecules that can play a nutritional role for the plant, which can be emphasized more due to the plant-based nature of Z. multiflora extract. In addition, it was observed during this study that nano-extract had a better effect on O. basilicum L.var. Genoves growth indices than regular Z. multiflora extract. Numerous studies have shown that the use of nanoparticles, by increasing the expression of plasma membrane aquaporins, plays an important role in root hydraulic conductivity and therefore plays an effective role in improving water absorption and reducing the effects of stress on the cell membrane [72].
The observed decrease in chlorophyll content due to salt stress in O. basilicum L. var. Genovese plants can be attributed to impaired absorption of mineral elements effective in chlorophyll synthesis such as nitrogen and iron, chlorophyll degradation or increased chlorophyllase enzyme activity, decreased chlorophyll synthase, production of reactive oxygen species, and destruction of photosynthetic pigments under stress conditions [67]. Salt stress disrupts ion absorption, which affects chloroplast growth and protein translation pathways in plastids. In the present study, a decrease in chlorophyll was observed under salinity conditions, and an increase in chlorophyll content was observed with the application of extract and nano extract of Z. multiflora compared to treatments without extract. Nanoextract may lead to an increased energy transfer from PSII to PSI with changes in the fluidity of the thylakoid membrane and improve the process of photosynthesis [73]. Extracts and nanoextracts may contain amino groups that maximize photosynthesis and ultimately increase plant growth [74].
Sodium ions, when entering the cytoplasm, have a strong inhibitory effect on the activity of many enzymes. One mechanism for reducing cellular sodium accumulation is its accumulation in the vacuole. The presence of sodium in the vacuole not only prevents contact with the cytosol but also maintains the osmotic balance inside the cell with its outside. The low concentration of cytosolic sodium and the balance of the potassium-to-sodium ratio [75]. Maintaining the sodium-to-potassium ratio in plant tissues is essential for regulating cell osmosis, maintaining swelling pressure, the activity of many enzymes, protein synthesis, oxidant metabolism, photosynthesis, and the opening and closing of stomata [76]. One of the possible mechanisms of Z. multiflora herbal extract for increasing stress tolerance in plants is increasing potassium, which in this way maintains the ionic balance in the plant and helps the plant absorb water under stress conditions. It can be concluded that the application of extracts and nano-extracts in plants under salinity stress, due to its effect on increasing the absorption and transport of minerals such as nitrogen, phosphorus, and potassium and maintaining ion homeostasis, causes the continuation of plant metabolism under stress conditions [77]. By using nano-fertilizers, the time and rate of element release are matched and coordinated with the plant’s nutritional needs. Therefore, the plant can absorb the maximum amount of nutrients and, while reducing the leaching of elements, the crop yield is also increased.
Flavonoids are among the most important secondary metabolites in plants that have strong antioxidant properties and their increase under stress conditions is a plant defense response to control free radicals produced under these conditions. The phenylpropanoid pathway is responsible for the synthesis of some phenolic metabolites in plants, most of which are produced under stress and have common precursors. Salinity stress reduces electron flow in the photosynthetic electron transport system and causes oxidative stress through the production of reactive oxygen species (ROS) [78]. To counteract toxic ROS, plants produce phenolic acids, flavonoids, and proanthocyanidins. Several studies in medicinal plants have shown that phenolics accumulate during salinity stress [79]. To protect cellular structures from the negative effects of salinity by scavenging free radicals and reducing oxidative damage. These compounds increase the stability of the cell wall and create a physical barrier against salinity stress [80]. Phenolic compounds reduce ROS through hydroxyl groups and also through metal-removing agents [81]. It is possible that foliar spraying of extracts and nano-extracts as elicitors may stimulate the synthesis of phenolic compounds by increasing the activity of the enzyme phenylalanine ammonia-lyase.
To reduce the negative effects of excess salt, plants synthesize and accumulate cytoplasmic osmolytes, including endogenous amino acids. The accumulation of proline in response to adverse environmental factors allows plants to adapt to stressful conditions, including salinity [82]. It can be said that in response to salinity stress, osmoregulation is a physiological adaptation that can be induced by the synthesis of compatible solutes or osmolytes such as proline, glycine betaine, amino acids, and soluble sugars to reduce the osmotic potential. The main function of osmolytes is to maintain cell turgor and maximize water uptake [83]. Proline acts as a cellular osmoprotectant and molecular chaperone to regulate osmosis, inhibit ROS, and stabilize macromolecules, DNA, membranes, and cells. Soluble sugar structures enhance proline biosynthesis and the antioxidant function of enzymes, leading to stress reduction [84]. In the present study, salinity increased proline accumulation. Excessive accumulation of compatible salts increases water uptake by increasing osmotic pressure, which maintains cell volume and positively affects physiological processes such as photosynthesis and stomatal conduction. It also enhances plant growth and tolerance to salinity stress [85].
Salinity tolerance is associated with the stimulation of antioxidant enzymes and their increased ability to remove reactive oxygen species. Catalase enzyme decomposes hydrogen peroxide without the need for energy, but this enzyme is active only at high concentrations of hydrogen peroxide, and its low concentrations are removed by peroxidase and peroxidases cooperating with strong reducing agents such as glutathione and ascorbate [86]. PAL enzyme is a key and determining enzyme at the beginning of the phenylpropanoid biosynthesis pathway and by deamination of the amino acid phenylalanine it converts it into trans-cinnamic acid, which is the first and most important intermediate for the production of phenolic compounds in plants [87]. The results of the present study showed that salinity stress generally increased the activity of these enzymes. Increased activity of PAL and other enzymes of the phenylpropanoid pathway leads to an increase in secondary metabolites in plants. The increase in PAL activity observed in the study showed that the plant’s defense mechanism has increased and the activities of this enzyme actually help the O. basilicum L.var. Genoves plants tolerate salt stress conditions. Due to the high mobility of nanoparticles, their transport to all parts of the plant is rapid. In addition, their small size and high surface area to volume ratio make them more accessible. In the present study, the superior effect of nanoextract was also observed.
The decrease in methyl chavicol simultaneously with the increase in some compounds (such as linalool) may indicate a change in biosynthetic pathways under stress. In this regard, it can be said that the severe decrease in methyl chavicol at 25-50 mM salinity (to 2.1% and 1.72%, respectively) may be due to the destruction of key enzymes of the phenylpropanoid pathway [88]. Also, the decrease in these compounds in salinity (especially alpha-humulene to 1.36%) may be due to the inhibition of the geranyl diphosphate-based terpenoid pathway [89]. Studies show that sesquiterpenes (such as alpha-humulene) are often reduced under severe stresses, as the plant diverts resources to less costly compounds (such as menthol) [90]. The sustained increase in linalool and menthol with increasing salinity (up to 23.91% and 13.62%) is consistent with studies showing that monoterpenes accumulate as defense compounds under stress conditions [91]. This is because these compounds help the plant cope with salt stress by activating antioxidant pathways (such as increasing catalase activity). The increase in eugenol up to 50 mM salinity (7.96%) and then decreasing is similar to reports indicating activation of the phenylpropanoid pathway at moderate salinity levels [92]. The decrease at 100 mM salinity is likely due to the degradation of the eugenol synthase enzyme under oxidative stress.
Environmental conditions significantly affect the composition of O. basilicum L. var. Genovese essential oil. Previous studies have shown that factors such as climate, geographical location, and soil structure and texture affect the yield of the plant’s essential oil [93,94]. The production of plant secondary metabolites is influenced by environmental conditions, especially biotic and abiotic stresses, among which increased salinity has a significant impact on plant essential oil biosynthesis and affects its composition [95]; therefore, in explaining the increase in essential oil content under stress conditions, it can be said that since the amount of primary metabolites in the plant decreases under adverse environmental conditions, the production of secondary metabolites increases as a defense mechanism to prevent intracellular oxidation in the plant [96].
5. Conclusions
The results showed that a significant decrease occurred in the growth and biochemical factors of O. basilicum L.var. Genoves under saline stress. According to the results obtained in this experiment, the highest efficiency under saline stress conditions was observed in the treatment of regular and nano extracts of Z. multiflora. O. basilicum L. var. Genoves plants can record higher or similar values of growth and physiological factors compared to control plants (without treatment) in moderate and severe salinity stresses. In general, the use of Z. multiflora extract and nano extract can be beneficial for inducing resistance in O. basilicum L. var. Genes of plants to salinity stress, and improving growth and increasing medicinal value.
Author Contributions
Conceptualization, E. Shabani. and F. Ghanbari.; methodology, A. Azizi.; software, A. Azizi. and E. Helalipour; validation, E. Shabani. and F. Ghanbari.; formal analysis, E. Helalipour; investigation, E. Shabani. and A. Azizi; resources, E. Helalipour; data curation, A. Azizi; writing—original draft preparation E. Shabani; A. Azizi. and E. Helalipour; writing—review and editing, E. Shabani. and M. Caser; visualization, A. Azizi; supervision, E. Shabani; project administration, E. Shabani. and F. Ghanbari; funding acquisition, E. Shabani. and F. Ghanbari. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
According to the rules of the university, the submission of thesis data for paper publishers and other universities is prohibited. So the data is confidential.
Acknowledgments
We would like to acknowledge vice chancellor for research at Shahid Chamran University of Ahvaz and Ilam University.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| PAL | Phenol Alanine Ammonia Lyase |
| ROS | Reactive Oxygen Species |
| CAT | Catalase enzyme |
| MDA | Malondialdehyde |
References
- Formisano, L.; Ciriello, M.; E.l-Nakhel, C.; Kyriacou, M.C.; Rouphael, Y. Successive harvests modulate the productive and physiological behavior of three genovese pesto basil cultivars. Agronomy. 2021, 11, 560. [CrossRef]
- Makri, O.; Kintzios, S. Ocimum sp. (basil): Botany, cultivation, pharmaceutical properties, and biotechnology. J. Herbs Spices Med. Plants. 2008, 13, 123–150. [Google Scholar] [CrossRef]
- Onofrei, V.; Benchennouf, A.; Jancheva, M.; Loupassaki, S.; Ouaret, W.; Burducea, M.; Lobiuc, A.; Teliban, G.C.; Robu, T. Ecological foliar fertilization effects on essential oil composition of sweet basil (Ocimum basilicum L.) cultivated in a field system. Sci. Hortic. 2018, 239, 104–113. [Google Scholar] [CrossRef]
- Shabani, E.; Bolandnazar, S.; Tabatabaei, S.J. Magnetized nutrient solution and arbuscular mycorrhizal affect essential oil and physiological aspects of sweet basil (Ocimum basilicum L.) grown in various P concentrations. J. Plant Nutr. 2022, 45, 883–895. [Google Scholar] [CrossRef]
- Jakovljević, D.; Momčilović, J.; Bojović, B.; Stanković, M. The Short-Term Metabolic Modulation of Basil (Ocimum basilicum L. cv.’Genovese’) after Exposure to Cold or Heat. Plants. 2021, 10, 590. [Google Scholar] [CrossRef] [PubMed]
- Dudai, N.; Belanger, F.C. Aroma as a factor in the Breeding Process of Fresh Herbs---The case of basil. Biotechnology in Flavor Production. 2009, 161–184. [Google Scholar] [CrossRef]
- Bahcesular, B.; Yildirim, E.D.; Karaçocuk, M.; Kulak, M.; Karaman, S. Seed priming with melatonin effects on growth, essential oil compounds and antioxidant activity of basil (Ocimum basilicum L.) under salinity stress. Ind Crops Prod. 2020, 146, 112165. [CrossRef]
- Maggio, A.; Roscigno, G.; Bruno, M.; De Falco, E.; Senatore, F. Essential-Oil Variability in a Collection of Ocimum basilicum L. Cultivars. Chem Biodivers. 2016, 13, 1357–1368. [Google Scholar] [CrossRef]
- Ciriello, M.; Formisano, L.; El-Nakhel, C.; Kyriacou, M.C.; Soteriou, G.A.; Pizzolongo, F.; Romano, R.; De Pascale, S.; Rouphael, Y. Genotype and successive harvests interaction affects phenolic acids and aroma profile of genovese basil for pesto sauce production. Foods. 2021, 10, 278. [Google Scholar] [CrossRef]
- Trettel, J.R.; Nascimento, A.B.; Barbosa, L.N.; Magalhaes, H.M. ‘In vitro’organogenesis and growth of’ Ocimum basilicum’’Genovese’(basil) cultivated with growth regulators. Aust. J. Crop Sci. 2019, 13, 1131–1140. [Google Scholar] [CrossRef]
- Bauer, K.; Garbe, D.; Surburg, H. Common fragrance and flavor materials: Preparation, properties and uses. John Wiley and Sons, 2008.
- Park, M.J.; Gwak, K.S.; Yang, I.; Kim, K.W.; Jeung, E.B.; Chang, J.W.; Choi, I.G. ; Effect of citral, eugenol, nerolidol and α-terpineol on the ultrastructural changes of Trichophyton rubrum. Fitoterapia. 2012, 83, 410–416. [Google Scholar] [CrossRef]
- Saia, S.; Corrado, G.; Vitaglione, P.; Colla, G.; Bonini, P.; Giordano, M.; Fiorillo, A.; Rouphael, Y. An Endophytic Fungi-Based Biostimulant Modulates Volatile and Non-Volatile Secondary Metabolites and Yield of Greenhouse Basil (Ocimum basilicum L.) through Variable Mechanisms Dependent on Salinity Stress Level. Pathogens. 2021, 10, 797. [Google Scholar] [CrossRef]
- Kanber, R.; Elhindi, K.M.; Alotaibi, M.A. Silicon supplementation mitigates salinity stress on Ocimum basilicum L. via improving water balance, ion homeostasis, and antioxidant defense system. cotoxicol. Environ. Saf 2020, 206, 111396. [Google Scholar] [CrossRef]
- .
- Kulak, M.; Gul, F.; Sekeroglu, N. Changes in growth parameter and essential oil composition of sage (Salvia officinalis L.) leaves in response to various salt stresses. Ind Crops Prod. 2020, 145, 112078. [Google Scholar] [CrossRef]
- Dias, N.S.; Fernandes, C.S.; Sousa-Neto, O.N.; Silva, C.R.; Ferreira, J.F.S.; Sa, F.V.S.; Cosme, C.R.; Souza, A.C.M.S.; Oliveira, A.M.; Batista, C.N.O. Potential agricultural use of reject brine from desalination plants in family farming areas. Saline and Alkaline Soils in Latin America. Natural Resources, Management and Productive Alternatives; 2021, 101–118. [CrossRef]
- Ghaemi, A.A.; Salimi, M.H.; Tabarza, A. The interaction of fishery effluent and plant residues on the yield and water consumption efficiency of cherry tomatoes under drip irrigation system in the greenhouse. J. Plant Interact. 2016, 8, 41–49. [Google Scholar] [CrossRef]
- Alexopoulos, A.A.; Marandos, E.; Assimakopoulou, A.; Vidalis, N.; Petropoulos, S.A.; Karapanos, I.C. Effect of nutrient solution pH on the growth, yield, and quality of Taraxacum officinale and Reichardia picroides in a floating hydroponic system. Agronomy. 2021, 11, 1118. [Google Scholar] [CrossRef]
- Sardare, M.D.; Admane, S.A. Review on plant without soil-hydroponics. IJRET. 2013, 02, 299–304. [Google Scholar]
- Liu, J.; Wang, F.; Liu, W.; Tang, C.; Wu, C.; Wu, Y. Nutrient removal by up-scaling a hybrid floating treatment bed (HFTB) using plant and periphyton: From laboratory tank to polluted river. Bioresour. Technol. 2016, 207, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H. Nitrogen removal during the cold season by constructed floating wetlands planted with Oenanthe javanica. Mar. Freshw. Res. 2018, 69, 635–647. [Google Scholar] [CrossRef]
- Ge, Z. .G.; Feng, C.M.; Wang, X.P.; Zhang, J.B. Seasonal applicability of three vegetation constructed floating treatment wetlands for nutrient removal and harvesting strategy in urban stormwater retention ponds. INT BIODETER BIODEGR. 2016, 112, 80–87. [Google Scholar] [CrossRef]
- Headley, T.R.; Tanner, C.C. Constructed wetlands with floating emergent macrophytes: An innovative stormwater treatment technology. rit Rev Environ Sci Technol. 2012, 42, 2261–2310. [Google Scholar] [CrossRef]
- Giménez, A.; Fernández, J.A.; Pascual, J.A.; Ros, M.; López-Serrano, M.; Egea-Gilabert, C. An agroindustrial compost as alternative to peat for production of baby leaf red lettuce in a floating system. Scientia Horticulturae. 2019, 246, 907–915. [Google Scholar] [CrossRef]
- Khater, E.S.; Bahnasawy, A.; Abass, W.; Morsy, O.; El-Ghobashy, H.; Shaban, Y.; Egela, M. Production of basil (Ocimum basilicum L.) under different soilless cultures. Adv. Environ. Sci. Technol. 2021, 11, 12754. [Google Scholar] [CrossRef]
- Moncada, A.; Miceli, A.; Vetrano, F. Use of plant growth-promoting rhizobacteria (PGPR) and organic fertilization for soilless cultivation of basil. Sci. Hortic. 2021, 275, 109733. [Google Scholar] [CrossRef]
- Rakocy, J.; Shultz, R.C.; Bailey, D.S.; Thoman, E.S. Aquaponic production of tilapia and basil: Comparing a batch and staggered cropping system. SPSCC. 2003, 648, 63–69. [Google Scholar] [CrossRef]
- Stoia, M.; Oancea, S. Low-molecular-weight synthetic antioxidants: Classification, pharmacological profile, effectiveness and trends. Antioxidants. 2022, 11, 638. [Google Scholar] [CrossRef]
- Rathee, P.; Sehrawat, R.; Rathee, P.; Khatkar, A.; Akkol, E.K.; Khatkar, S.; Sobarzo-Sánchez, E. Polyphenols: Natural preservatives with promising applications in food, cosmetics and pharma industries; problems and toxicity associated with synthetic preservatives; impact of misleading advertisements; recent trends in preservation and legislation. Materials. 2023, 16, 4793. [Google Scholar] [CrossRef] [PubMed]
- Rathee-Gomes, E.A.; del, C.; Mejia-da-Silva, L.; Pérez-Mora, L.S.; Matsudo, M.C.; Ferreira-Camargo, L.S.; Singh, A.K.; de Carvalho, J.C.M. Potential of microalgae carotenoids for industrial application. Appl. Biochem. Biotechnol. 2019, 188, 602–634. [Google Scholar] [CrossRef] [PubMed]
- Shahid, I.; Mehnaz, S. Microbial secondary metabolites: Effectual armors to improve stress survivability in crop plants. In Microbial Services in Restoration Ecology; 2020, pp. 47–70. [CrossRef]
- Dutta, S.; Ray, S. Comparative assessment of total phenolic content and in vitro antioxidant activities of bark and leaf methanolic extracts of Manilkara hexandra (Roxb.) Dubard. J. King Saud Univ. Sci. 2020, 32, 643–647. [Google Scholar] [CrossRef]
- Agarwal, S.; Pandey, V. Antioxidant enzyme responses to NaCl stress in Cassia angustifolia. Biol. Plant. 2004, 48, 555–560. [Google Scholar] [CrossRef]
- Kumaran, A. Antioxidant and free radical scavenging activity of an aqueous extract of Coleus aromaticus. Food Chem. 2006, 97, 109–114. [Google Scholar] [CrossRef]
- Stoilova, I.; Krastanov, A.; Stoyanova, A.; Denev, P.; Gargova, S. Antioxidant activity of a ginger extract (Zingiber officinale). Food Chem. 2007, 102, 764–770. [Google Scholar] [CrossRef]
- Ali, E.; Hassan, F.; Elgimabi, M. Improving the growth, yield and volatile oil content of Pelargonium graveolens L. Herit by foliar application with moringa leaf extract through motivating physiological and biochemical parameters. S Afr J Bot. 2018, 119, 383–389. [Google Scholar] [CrossRef]
- Diab, F.; Khalil, M.; Lupidi, G.; Zbeeb, H.; Salis, A.; Damonte, G.; Bramucci, M.; Portincasa, P.; Vergani, L. Influence of simulated in vitro gastrointestinal digestion on the phenolic profile, antioxidant, and biological activity of Thymbra Spicata L. extracts. Antioxidants. 2022, 11, 1778. [Google Scholar] [CrossRef] [PubMed]
- Solgi, M.; Bagnazari, M.; Mohammadi, M.; Azizi, A. Thymbra spicata extract and arbuscular mycorrhizae improved the morphophysiological traits, biochemical properties, and essential oil content and composition of Rosemary (Rosmarinus officinalis L.) under salinity stress. BMC Plant Biol. 2025, 25, 220. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, S.Y. Antioxidant activity and phenolic compounds in selected herbs. JAFC. 2001, 49, 5165–5170. [Google Scholar] [CrossRef] [PubMed]
- Bonesi, M.; Loizzo, M.R.; Acquaviva, R.; Malfa, G.A.; Aiell, F.; Tundis, R. Anti-inflammatory and antioxidant agents from Salvia genus (Lamiaceae): An assessment of the current state of knowledge. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2017, 16, 70–86. [CrossRef]
- Bonesi, S.; Pimoradloo, E.; Bonesi, M.; Vessal, M. Evaluation of antioxidant potentials and α-amylase inhibition of different fractions of labiatae plants extracts: As a model of antidiabetic compounds properties. BioMed Research International. 2017, 2017, 7319507. [Google Scholar] [CrossRef]
- Mimica-Dukic, N.; Bozin, B. Mentha L. species (Lamiaceae) as promising sources of bioactive secondary metabolites. Current Pharmaceutical Design. 2008, 14, 3141–3150. [Google Scholar] [CrossRef]
- Ahari, H.; Yousefi, S. Green synthesis of three-component Ag/AgCl/TiO2 nanocomposite using Zataria Multiflora plant. Journal of food science and technology (Iran). 2023, 20, 94–112. [Google Scholar] [CrossRef]
- Golkar, P.; Mosavat, N.; Jalali, S.A.H. Essential oils, chemical constituents, antioxidant, antibacterial and in vitro cytotoxic activity of different Thymus species and Zataria multiflora collected from Iran. S Afr J Bot. 2020, 130, 250–258. [Google Scholar] [CrossRef]
- Akrami, F.; Rodríguez-Lafuente, A.; Bentayeb, K.; Pezo, D.; Ghalebi, S.R.; Nerín, C. Antioxidant and antimicrobial active paper based on Zataria (Zataria multiflora) and two cumin cultivars (Cuminum cyminum). LWT - Food Sci. Technol. 2015, 60, 929–933. [Google Scholar] [CrossRef]
- Pandey, G. Agri-Nanotechnology for sustainable agriculture, in: J. Sustain. Agric. 2020, 229–249. [Google Scholar] [CrossRef]
- Hofmann, T.; Lowry, G.V.; Ghoshal, S.; Tufenkji, N.; Brambilla, D.; Dutcher, J.R.; Wilkinson, K.J. Technology readiness and overcoming barriers to sustainably implement nanotechnology-enabled plant agriculture. Nat. Food. 2020, 1, 416–425. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Srivastava, A.K.; Elsadek, M.S.A.; Kordrostami, M.; Tran, L.S.P. Titanium dioxide nanoparticles improve growth and enhance the tolerance of broad bean plants under saline soil conditions. LAND DEGRAD DEV. 2018, 29, 1065–1073. [Google Scholar] [CrossRef]
- Soleymanzadeh, R.; Iranbakhsh, A.; Habibi, G.; Ardebili, Z.O. Selenium nanoparticle protected strawberry against salt stress through modifications in salicylic acid, ion homeostasis, antioxidant machinery, and photosynthesis performance. ACTA BIOL CRACOV BOT. 2020, 62, 33–42. [Google Scholar] [CrossRef]
- Arora, S.; Sharma, P.; Kumar, S.; Nayan, R.; Khanna, P.K.; Zaidi, M.G.H. Gold-nanoparticle induced enhancement in growth and seed yield of Brassica juncea. Plant Growth Regul. 2012, 66, 303–310. [Google Scholar] [CrossRef]
- Manke, A.; Wang, L.; Rojanasakul, Y. Mechanisms of nanoparticle-induced oxidative stress and toxicity. Biomed Res. Int. 2013, 2013, 942916. [Google Scholar] [CrossRef]
- Caser, M.; Lovisolo, C.; Scariot, V. The influence of water stress on growth, ecophysiology and ornamental quality of potted Primula vulgaris ‘Heidy’ plants. New insights to increase water use efficiency in plant production. J. Plant Biochem Physiol. 2024, 196, 111–122. [Google Scholar] [CrossRef]
- El-Berawey, D.Y.; Eldebawy, E.M.M. The effects of Marrubium alysson and Torilis arvensis natural and nano extracts on priming of wheat seeds in response to drought. Cereal Res. Commun. 2025, 53, 275–289. [Google Scholar] [CrossRef]
- Ghazy, O.A.; Fouad, M.T.; Saleh, H.H.; Kholif, A.E.; Morsy, T.A. Ultrasound-assisted preparation of anise extract nanoemulsion and its bioactivity against different pathogenic bacteria. Food Chem. 2021, 341, 128259. [Google Scholar] [CrossRef]
- Arnon, A.N. Method of extraction of chlorophyll in the plants. Agronomy journal. 1967, 23, 112–121. [Google Scholar]
- Pandjaitan, N.; Howard, L.R.; Morelock, T.; Gil, M.I. Antioxidant capacity and phenolic content of spinach as affected by genetics and maturation. J Agric Food Chem. 2005, 53, 8618–8623. [Google Scholar] [CrossRef]
- Krizek, D.T.; Britz, S.J.; Mirecki, R.M. Inhibitory effects of ambient levels of solar UV-A and UV-B radiation on growth of cv. New Red Fire lettuce. Physiol. Plant. 1998, 103, 1–7. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.A.; Teare, I.D. Rapid determination of free proline for water-stress studies. J Plant Nutr Soil Sci. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. METHOD ENZYMOL. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, L.; W.u, J.; Tan, R. Involvement of nitric oxide in oxidative burst, phenylalanine ammonia-lyase activation and Taxol production induced by low-energy ultrasound in Taxus yunnanensis cell suspension cultures. Nitric Oxide. 2006, 15, 351–358. [CrossRef]
- Akowuah, G.A.; Ismail, Z.; Norhayati, I.; Sadikun, A. The effects of different extraction solvents of varying polarities on polyphenols of Orthosiphon stamineus and evaluation of the free radical-scavenging activity. Food Chem. 2005, 93, 311–317. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Analytical biochemistry. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Tandon, H.L.S. Methods of analysis of soils, plants, water andfertilizers. FDCO, New Delhi; 1995.
- Adams, R.P. Identification of essential oil components by gas chromatography/mass spectrometry, ed. 4.1. Allured publishing corporation; 2007.
- Pirbalouti, A.G.; Hashemi, M.; Ghahfarokhi, F.T. Essential oil and chemical compositions of wild and cultivated Thymus daenensis Celak and Thymus vulgaris L. Ind Crops Prod. 2013, 48, 43–48. [Google Scholar] [CrossRef]
- Rezende, R.A.L.S.; Rodrigues, F.A.; Soares, J.D.R.; Silveira, H.R.O.D.; Pasqual, M.; Dias, G.M.G. Nitrogen removal during the cold season by constructed floating wetlands planted with Oenanthe javanica. Mar. Freshw. Res. 2018, 69, 635–647. [Google Scholar] [CrossRef]
- Abdel-Latif, A.; El-Demerdash, F.M. The ameliorative effects of silicon on salt-stressed sorghum seedlings and its influence on the activities of sucrose synthase and PEP carboxylase. Physiol. Plant Pathol. 2017, 5, 2–8. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Marashi, A.M.; Falah, N.A. Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. J. Chem. Health Risks. 2014, 4, 49–55. [Google Scholar]
- Chehregani Rad, A.; Khorzaman, N.; LariYazdi, H.; Shirkhani, Z. Changes in growth characteristics and physiological indices in Zn-Stressed Phaseolus vulgaris plants on hydroponic medium. J. Dev. Biol. 2016, 8, 31–39. [Google Scholar]
- Mahlooji, M. Effects of salinity stress and Zinc application and some physiological traits on grain filling of three barley cultivars. Journal of Plant Process and Function. 2022, 11, 211–227. [Google Scholar]
- Zulfiqar, F.; Ashraf, M. Nanoparticles potentially mediate salt stress tolerance in plants. Plant Physiol. Biochem. 2021, 160, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Hniličková, H.; Hnilička, F.; Martinkova, J.; Kraus, K. Effects of salt stress on water status, photosynthesis and chlorophyll fluorescence of rocket. Plant Soil Environ. 2017, 63, 362–367. [Google Scholar] [CrossRef]
- Sofy, M.R.; Elhawat, N.; Alshaal, T. Glycine betaine counters salinity stress by maintaining high K+/Na+ ratio and antioxidant defense via limiting Na+ uptake in common bean (Phaseolus vulgaris L.). Ecotoxicol. Environ. Saf. 2020, 200, 110732. [Google Scholar] [CrossRef]
- Summart, J.; Thanonkeo, P.; Panichajakul, S.; Prathepha, P.; McManus, M.T. Effect of salt stress on growth, inorganic ion and proline accumulation in Thai aromatic rice, Khao Dawk Mali 105, callus culture. Afr. J. Biotechnol. 2010, 9, 145–152. [Google Scholar]
- Shabala, S.; Demidchik, V.; Shabala, L.; Cuin, T.A.; Smith, S.J.; Miller, A.J.; Davies, J.M.; Newman, I.A. Extracellular Ca2+ ameliorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. J. Plant Physiol. 2006, 141, 1653–1665. [Google Scholar] [CrossRef]
- Carmona, F.J.; Guagliardi, A.; Masciocchi, N. Nanosized calcium phosphates as novel macronutrient nano-fertilizers. J. Nanomater. 2022, 12, 2709. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. environ. sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Waskiewicz, A.; Muzolf-Panek, M.; Golinski, P. Phenolic content changes in plants under salt stress. In: J. Plant Stress Physiol. 2013, 283–314. [CrossRef]
- Firoozeh, R.; Khavarinejad, R.; Najafi, F.; Saadatmand, S. Effects of gibberellin on contents of photosynthetic pigments, proline, phenol and flavonoid in savory plants (Satureja hortensis L.) under salt stress. Iran. J. Plant Physiol. 2019, 31, 894–908. [Google Scholar]
- Isah, T. Stress and defense responses in plant secondary metabolites production. J. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef]
- Amist, N.; Singh, N.B. Responses of enzymes involved in proline biosynthesis and degradation in wheat seedlings under stress. J. Biol. Res. 2017, 42, 195–206. [Google Scholar] [CrossRef]
- Hmidi, D.; Abdelly, C.; Athar, H.U.R.; Ashraf, M.; Messedi, D. Effect of salinity on osmotic adjustment, proline accumulation and possible role of ornithine-δ-ami notransferase in proline biosynthesis in Cakile maritima. Physiol. Mol. Biol. Plants. 2018, 24, 1017–1033. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, M.M.; Pradhan, N.; Subba, R.; Saha, P.; Roy, S. Sugar-terminated carbon-nanodots stimulate osmolyte accumulation and ROS detoxification for the alleviation of salinity stress in Vigna radiata. Sci. Rep. 2022, 12, 17567. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M. Interactive role of nitric oxide and calcium chloride in enhancing tolerance to salt stress. Nitric Oxide. 2012, 27, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.B.; Chu, L.Y.; Shao, M.A.; Jaleel, C.A.; Hong-mei, M. Higher plant antioxidants and redox signaling under environmental stresses. C R Biol. 2008, 331, 433–441. [Google Scholar] [CrossRef]
- Wen, P.F.; Chen, J.Y.; Wan, S.B.; Kong, W.F.; Zhang, P.; Wang, W.; Huang, W.D. Salicylic acid activates phenylalanine ammonia-lyase in grape berry in response to high temperature stress. J. Plant Growth Regul. 2008, 55, 1–10. [Google Scholar] [CrossRef]
- Rezaie, R.; Abdollahi Mandoulakani, B.; Fattahi, M. Cold stress changes antioxidant defense system, phenylpropanoid contents and expression of genes involved in their biosynthesis in Ocimum basilicum L. Sci. Rep. 2020, 10, 5290. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V.; Niazi, A.; Moghadam, A. Effect of salt stress on terpenoid biosynthesis in Salvia mirzayanii: From gene to metabolite. J. Hortic. Sci. Biotechnol. 2019, 94, 389–399. [Google Scholar] [CrossRef]
- El Amerany, F. The role of terpenoids in plant development and stress tolerance. In: Molecular and Physiological Insights into Plant Stress Tolerance and Applications in Agriculture—Part 2. Bentham Science Publishers; 2024, 71–98. [CrossRef]
- Selmar, D.; Kleinwächter, M. Stress enhances the synthesis of secondary plant products: The impact of stress-related over-reduction on the accumulation of natural products. Plant Cell Physiol. 2013, 54, 817–826. [Google Scholar] [CrossRef]
- Rafeie, M.; Shabani, L.; Sabzalian, M.R.; Gharibi, S. Pretreatment with LEDs regulates antioxidant capacity and polyphenolic profile in two genotypes of basil under salinity stress. Protoplasma. 2022, 259, 1567–1583. [Google Scholar] [CrossRef]
- Lo, Presti.; M., Ragusa.; S., Trozzi.; A., Dugo.; P., Visinoni.; F., Fazio.; A., Dugo.; G., Mondello.; L. A comparison between different techniques for the isolation of rosemary essential oil. J. Sep. Sci. 2005, 28, 273–280. [CrossRef]
- Gachkar, L.; Yadegari, D.; Rezaei, M.B.; Taghizadeh, M.; Astaneh, S.A.; Rasooli, I. Chemical and biological characteristics of Cuminum cyminum and Rosmarinus officinalis essential oils. Food Chem. 2007, 102, 898–904. [Google Scholar] [CrossRef]
- Karray-Bouraoui, N.; Rabhi, M.; Neffati, M.; Baldan, B.; Ranieri, A.; Marzouk, B.; Lachaal, M.; Smaoui, A. Salt effect on yield and composition of shoot essential oil and trichome morphology and density on leaves of Mentha pulegium. IND CROP PROD. 2009, 30, 338–343. [Google Scholar] [CrossRef]
- Bernstein, N.; Kravchik, M.; Dudai, N. Salinity-induced changes in essential oil, pigments and salts accumulation in sweet basil (Ocimum basilicum) in relation to alterations of morphological development. Ann. Appl. Biol. 2010, 156, 167–177. [Google Scholar] [CrossRef]
Figure 1.
TEM image of Z. multiflora extract nanoemulsion.
Figure 2.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on Total phenol (A), Flavonoids (B), and Proline (C). In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
Figure 2.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on Total phenol (A), Flavonoids (B), and Proline (C). In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
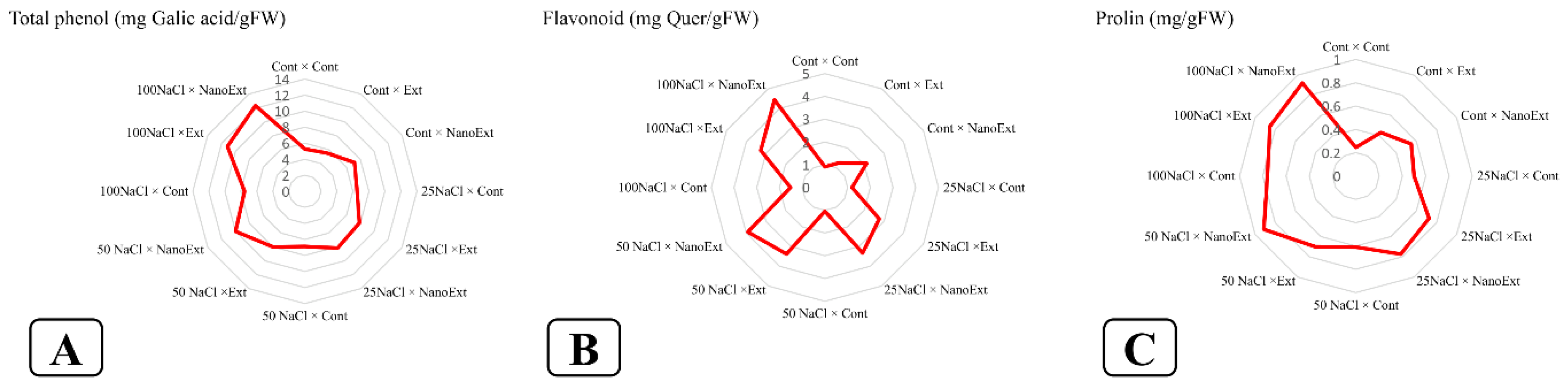
Figure 3.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on enzymes of Catalase (A), and Phenolalanine Ammonialyase (PAL) (B). In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
Figure 3.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on enzymes of Catalase (A), and Phenolalanine Ammonialyase (PAL) (B). In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
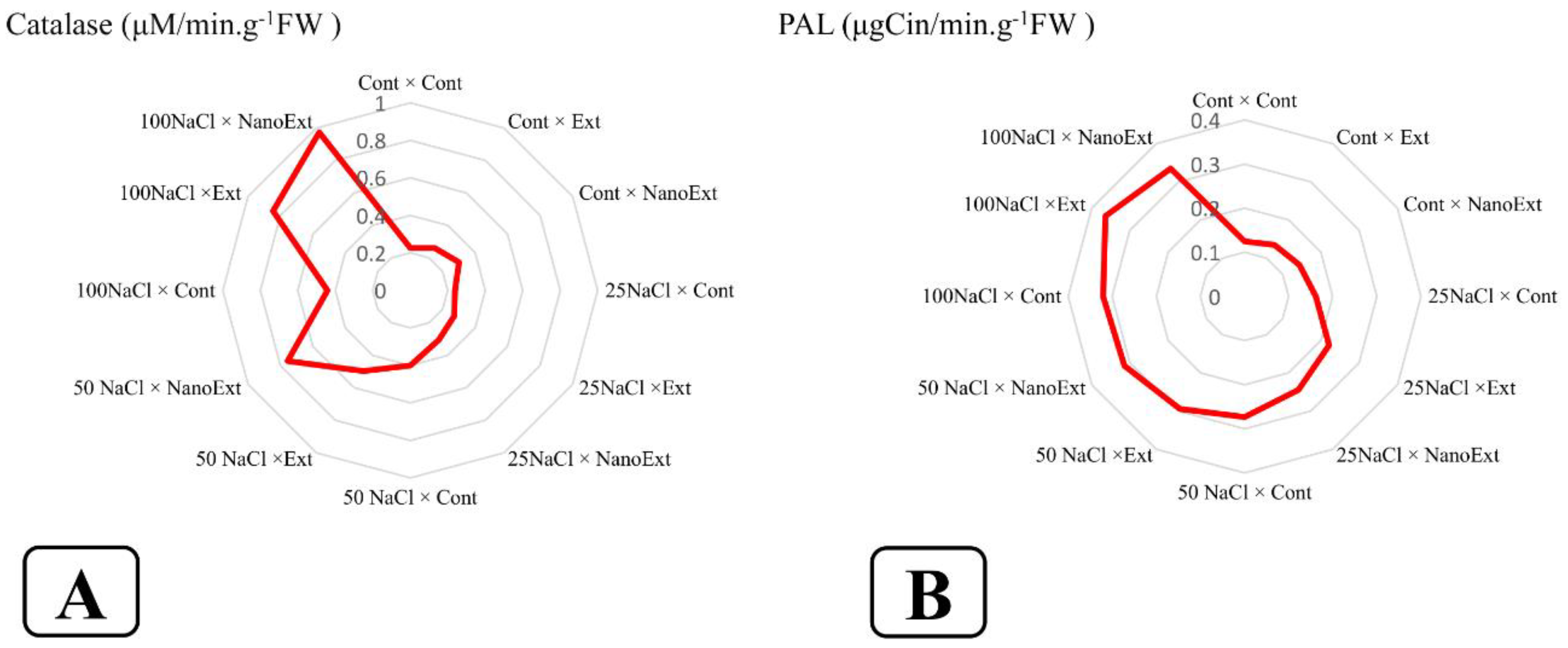
Figure 4.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on antioxidant activity, DPPH (A), and FRAP (B). In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
Figure 4.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on antioxidant activity, DPPH (A), and FRAP (B). In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
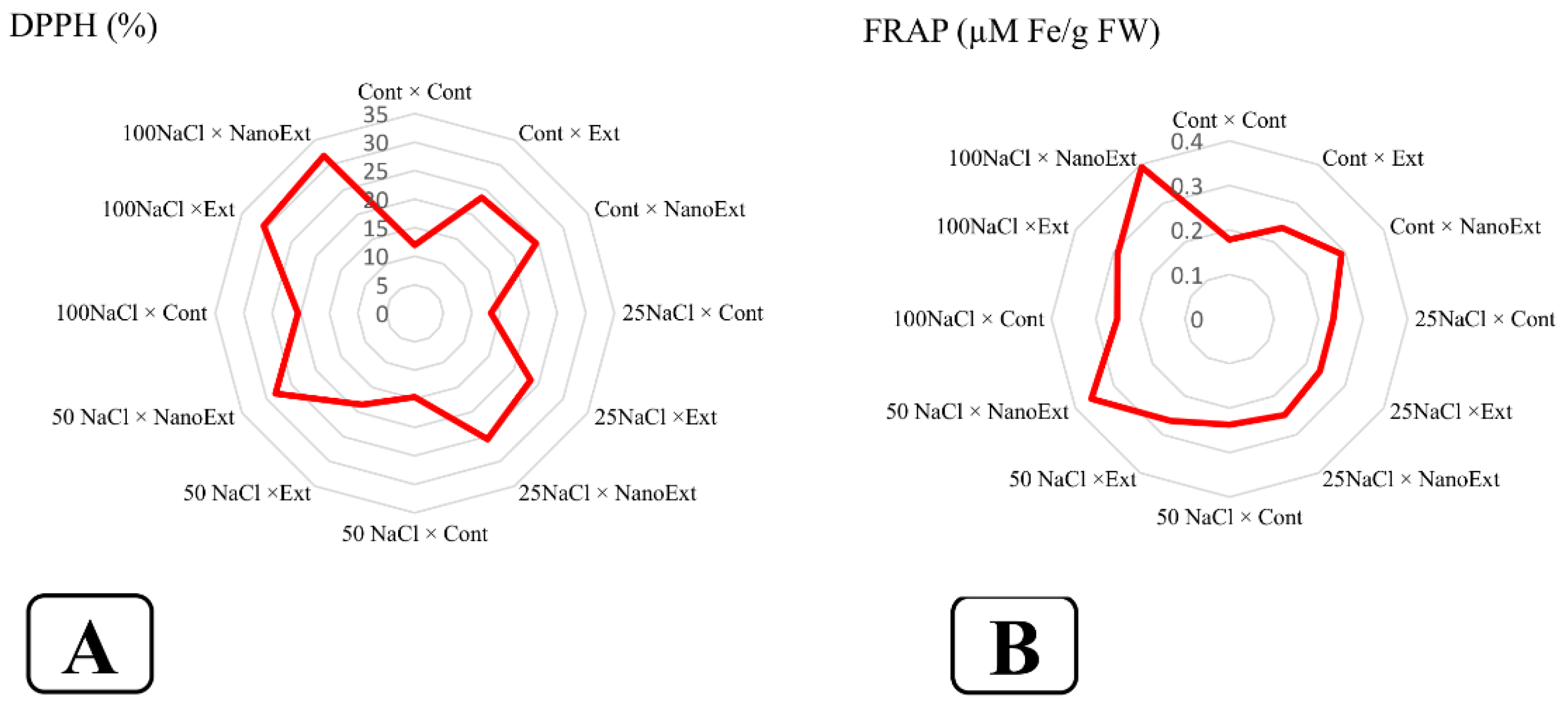
Figure 5.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on Methyl Chavicol content. In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
Figure 5.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on Methyl Chavicol content. In this figure, Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
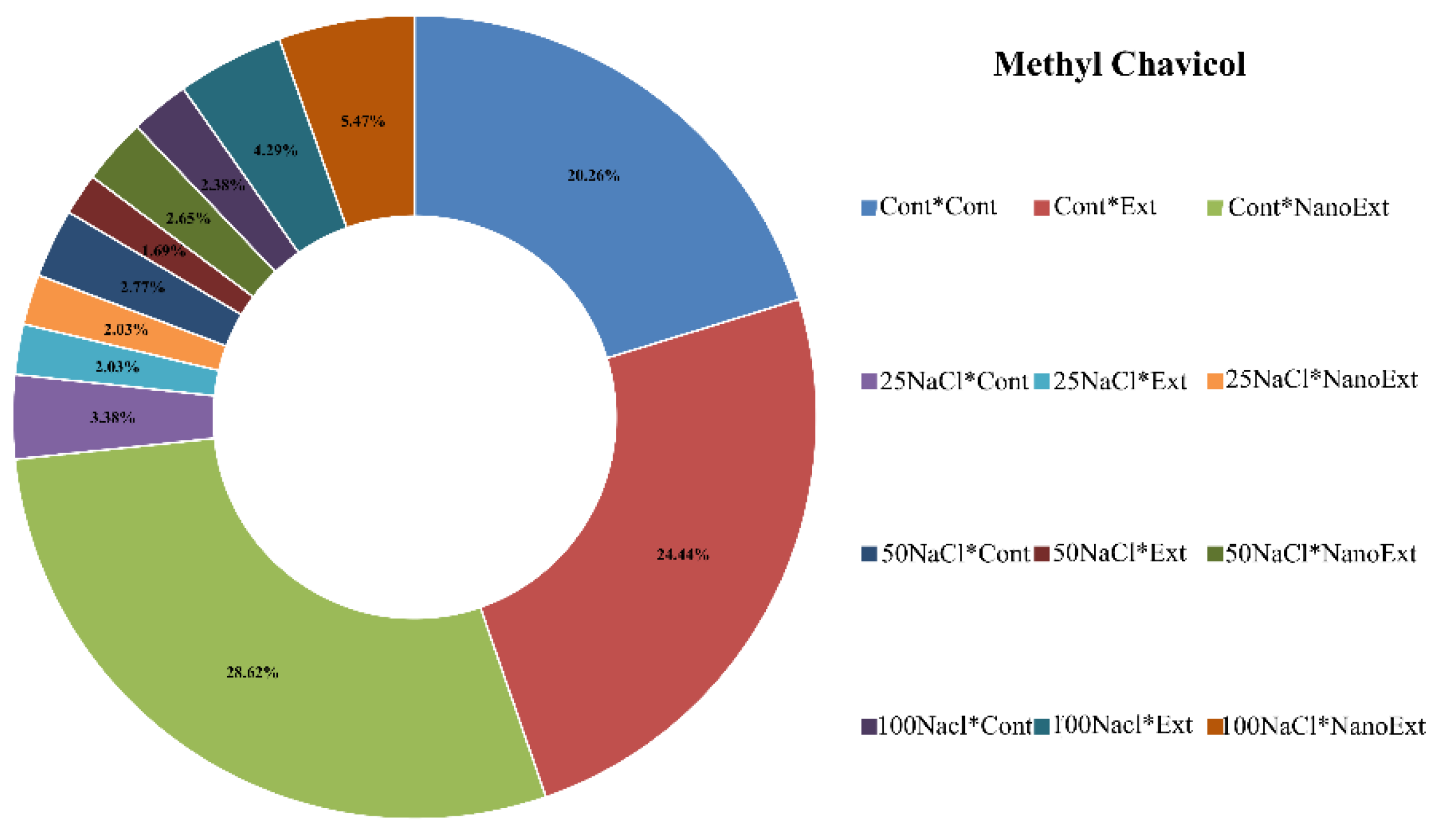
Figure 6.
Heatmap (A) and Bipolt (B) Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on morphophysiological and biochemical traits of O. basilicum L.var. Genoves (). In this figure, F.SH (shoot fresh weight), D.SH (shoot dry weight), F.R (root fresh weight), D.R (root dry weight), chl T (total chlorophyll), TPC (total phenol), TFC (flavonoid), CAT (catalase enzyme), PAL (phenylalanine ammonia-lyase enzyme), Antioxidant activity (DPPH), Antioxidant activity (FARP), PROL (proline), Na (sodium) and K (potassium) are shown.
Figure 6.
Heatmap (A) and Bipolt (B) Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on morphophysiological and biochemical traits of O. basilicum L.var. Genoves (). In this figure, F.SH (shoot fresh weight), D.SH (shoot dry weight), F.R (root fresh weight), D.R (root dry weight), chl T (total chlorophyll), TPC (total phenol), TFC (flavonoid), CAT (catalase enzyme), PAL (phenylalanine ammonia-lyase enzyme), Antioxidant activity (DPPH), Antioxidant activity (FARP), PROL (proline), Na (sodium) and K (potassium) are shown.
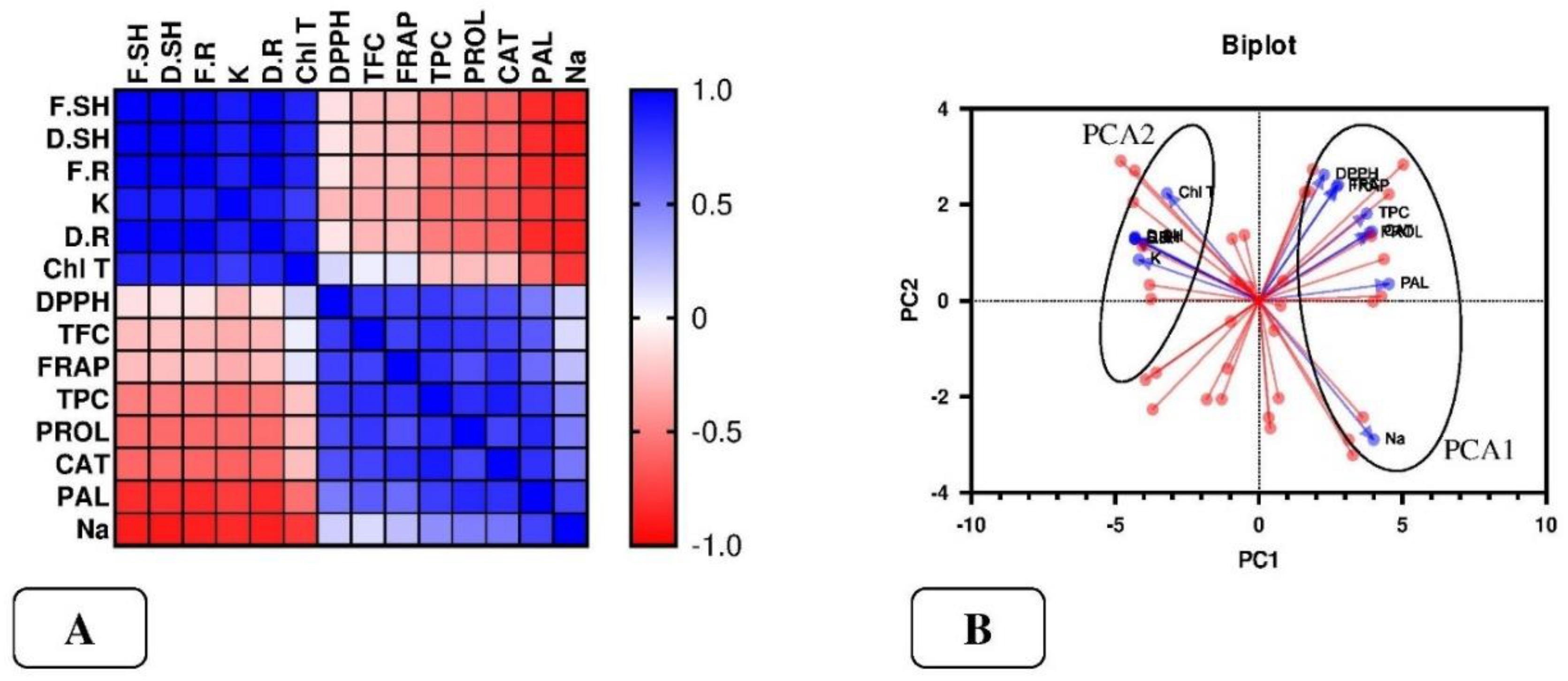

Table 1.
Comparison of the average effect of ethanol and nano-extract of Z. multiflora (Control (Cont), regular extract (Ext) and nano-extract (NanoExt)) on vegetative and physiologic traits of O. basilicum L. var. Genovese under floating culture in salinity condtion (Cont= 0 mM of NaCl salinity, 25NaCl= 25 mM of NaCl salinity, 50NaCl = 50 mM of NaCl salinity, 100NaCl = 100 mM of NaCl salinity).
Table 1.
Comparison of the average effect of ethanol and nano-extract of Z. multiflora (Control (Cont), regular extract (Ext) and nano-extract (NanoExt)) on vegetative and physiologic traits of O. basilicum L. var. Genovese under floating culture in salinity condtion (Cont= 0 mM of NaCl salinity, 25NaCl= 25 mM of NaCl salinity, 50NaCl = 50 mM of NaCl salinity, 100NaCl = 100 mM of NaCl salinity).
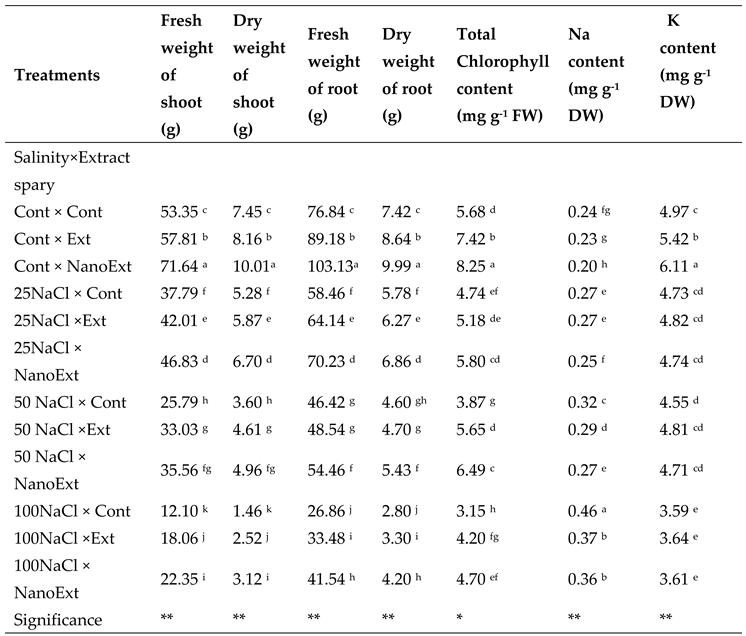 |
* p < 0.05; ** p < 0.01. Mean values with the same letter were not different according to the Duncan’s test (p < 0.05).
Table 2.
Analysis variance of the effect of Z. multiflora extract (Control, regular extract, and nano-extract) on bichemical traits of O. basilicum L. var. Genovese under floating culture in salinity condtion.
Table 2.
Analysis variance of the effect of Z. multiflora extract (Control, regular extract, and nano-extract) on bichemical traits of O. basilicum L. var. Genovese under floating culture in salinity condtion.
| Treatment | df | Phenol | Flavonoid | Proline | Catalase | Phenolalanine Ammonialyase | DPPH | FRAP |
|---|---|---|---|---|---|---|---|---|
| Block | 2 | 2.05 ** | 0.03 NS | 0.00 NS | 0.006 ** | 0.00 NS | 18.16 * | 0.002 * |
| Salinity (S) | 3 | 29.27 ** | 4.65 ** | 0.31 ** | 0.50 ** | 0.07 ** | 119.25 ** | 0.01 ** |
| Error a | 4 | 0.46 | 0.07 | 0.00 | 0.001 | 0.00 | 3.95 | 0.00 |
| Extract spray (E) | 2 | 11.26 ** | 10.07 ** | 0.19 ** | 0.06 ** | 0.005 ** | 378.77 ** | 0.01 ** |
| S×E | 4 | 1.23 ** | 0.86 ** | 0.007 ** | 0.02 ** | 0.00 * | 19.81 * | 0.002 * |
| Error b | 18 | 0.31 | 0.08 | 0.0002 | 0.00 | 0.00 | 4.68 | 0.00 |
| CV(%) | - | 6.93 | 11.90 | 2.38 | 0.02 | 0.01 | 9.74 | 10.01 |
* p < 0.05; ** p < 0.01; NS, not significant. Mean values with the same letter were not different according to the Duncan’s test (p < 0.05).
Table 3.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on Essential oil content. Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
Table 3.
Effect of extract and nano-extract of Z. multiflora on salt stress tolerance of O. basilicum L.var. Genoves under floating cultivation on Essential oil content. Cont (0 mM of NaCl salinity), 25NaCl (25 mM of NaCl salinity), 50NaCl (50 mM of NaCl salinity), 100NaCl (100 mM of NaCl salinity), and Cont (control, without extract spraying), Ext (foliar spray of regular extract of Z. multiflora), and NanoExt (foliar spray of nano extract of Z. multiflora).
| Cont × Cont | Cont × Ext | Cont × NanoExt | 25NaCl × Cont | 25NaCl ×Ext | 25NaCl × NanoExt | 50 NaCl × Cont | 50 NaCl ×Ext | 50 NaCl × NanoExt | 100NaCl × Cont | 100NaCl ×Ext | 100NaCl × NanoExt | ||
| Compound | KI | Area% | Area% | Area% | Area% | Area% | Area% | Area% | Area% | Area% | Area% | Area% | Area% |
| a-Pinene | 939 | 0.14 | 0.32 | 0.45 | 0.61 | 0.85 | 0.54 | 1.25 | 1.12 | 1.36 | 0.63 | 0.85 | 0.93 |
| Sabinene | 975 | 0.97 | 1.12 | 1.36 | 1.49 | 1.64 | 1.96 | 2.36 | 1.39 | 0.48 | 0.63 | 0.78 | 0.96 |
| Myrcene | 989 | 0.41 | 0.36 | 0.89 | 0.92 | 1.26 | 0.15 | 0.54 | 0.64 | 0.78 | 0.37 | 0.54 | 0.24 |
| 1,8-Cineol | 1030 | 1.52 | 1.45 | 2.58 | 3.26 | 2.31 | 2.13 | 3.79 | 2.15 | 1.65 | 2.98 | 2.45 | 2.47 |
| Linalool | 1088 | 12.3 | 13.25 | 15.3 | 22.64 | 18.26 | 21.26 | 22.25 | 23.13 | 19.94 | 21.47 | 22.68 | 23.91 |
| Eugenol | 1355 | 2.4 | 3.4 | 3.85 | 3.95 | 5.84 | 6.84 | 7.26 | 7.96 | 5.95 | 4.78 | 4.55 | 4.15 |
| Camphor | 1145 | 0.74 | 0.89 | 0.65 | 0.74 | 0.61 | 0.64 | 0.97 | 1.14 | 8.56 | 5.14 | 6.37 | 6.29 |
| Terpinen-4-ol | 1177 | 0.41 | 0.61 | 0.74 | 0.85 | 0.45 | 0.79 | 0.85 | 1.29 | 2.14 | 1.34 | 2.78 | 2.18 |
| a-Terpineol | 1188 | 1.54 | 1.2 | 2.3 | 3.2 | 3.28 | 2.15 | 1.02 | 2.46 | 3.26 | 2.96 | 3.79 | 2.65 |
| Cis-Carveol | 1229 | 0.65 | 0.64 | 0.78 | 0.64 | 0.79 | 0.75 | 0.63 | 0.98 | 1.24 | 2.78 | 2.36 | 2.06 |
| Geraniol | 1265 | 0.41 | 0.18 | 0.52 | 0.56 | 0.14 | 0.62 | 2.36 | 0 | 0 | 0 | 0 | 0 |
| Cubenol | 1511 | 2.7 | 1.36 | 1.25 | 1.39 | 2.31 | 1.12 | 0 | 0 | 0 | 0 | 0 | 0 |
| Epi-a-Muurolol | 1642 | 1.47 | 1.78 | 1.97 | 3.24 | 5.6 | 2.03 | 0 | 0 | 0 | 0 | 0 | 0 |
| E-B-Ocimene | 1045 | 2.1 | 1.2 | 2.5 | 2.6 | 3.1 | 1.46 | 1.25 | 2.15 | 1.02 | 3.98 | 3.94 | 4.18 |
| Carvone | 1243 | 15.9 | 16.6 | 16.4 | 18.23 | 9.24 | 12.4 | 14.26 | 13.6 | 7.6 | 7.63 | 7.37 | 7.7 |
| 6-Methyl-5-hepten-2-one | 989 | 0.35 | 0.21 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Methyl Chavicol | 1196 | 12.6 | 15.2 | 17.8 | 2.1 | 1.26 | 1.26 | 1.72 | 1.05 | 1.65 | 1.48 | 2.67 | 3.4 |
| Nero | 1230 | 0.62 | 0.85 | 1.25 | 1.24 | 2.12 | 1.02 | 2.36 | 3.65 | 2.15 | 1.96 | 1.64 | 1.85 |
| Neral | 1242 | 0.52 | 0.62 | 0.65 | 0.89 | 3.16 | 0.96 | 0.89 | 0.85 | 0.89 | 0.52 | 0.53 | 0.63 |
| A-Humulene | 1456 | 10.2 | 8.25 | 5.12 | 4.29 | 2.12 | 2.31 | 1.56 | 1.36 | 2.16 | 3.46 | 2.98 | 3.26 |
| Actanol acetate | 1213 | 0.5 | 0.52 | 0.62 | 0.89 | 1.21 | 0 | 0 | 0 | 0.39 | 0 | 0 | 0 |
| 1-Octen-3-ol | 980 | 0.35 | 0.41 | 0.23 | 0.27 | 0.36 | 0 | 0 | 0 | 0.46 | 0 | 0 | 0 |
| Geranial | 1272 | 6.2 | 0.74 | 0.84 | 0.82 | 0.8 | 1.96 | 1.36 | 1.96 | 0.52 | 0.45 | 0.63 | 0.59 |
| E-Caryophyllene | 1418 | 0.5 | 0.65 | 0.79 | 0.8 | 0.75 | 0.61 | 0.98 | 1.97 | 0.17 | 0.52 | 0.78 | 0.64 |
| Isopulego | 1150 | 0.41 | 0.12 | 0.15 | 0 | 0 | 0.45 | 0.45 | 0.89 | 0.61 | 0.61 | 0.96 | 0.79 |
| Italicence ether | 1537 | 0.65 | 0.87 | 1.25 | 1.24 | 1.25 | 2.89 | 1.26 | 1.52 | 0.36 | 0.89 | 1.02 | 1.36 |
| B-Pinene | 979 | 1.4 | 2.14 | 1.36 | 4.58 | 3.46 | 2.16 | 0.84 | 0.94 | 0.34 | 0.98 | 2.16 | 1.03 |
| Menthol | 1171 | 4.2 | 5.6 | 6.2 | 6.85 | 9.03 | 9.25 | 10.39 | 11.26 | 10.15 | 11.57 | 12.48 | 13.62 |
| DeltGuaie | 1025 | 0.95 | 0.89 | 0.65 | 0.45 | 0 | 0.45 | 0.65 | 0.64 | 0.12 | 1.36 | 1.65 | 2.49 |
| Cabbenol | 1136 | 1.14 | 2.15 | 0.46 | 0.35 | 0 | 0 | 0.45 | 0.35 | 0 | 0 | 0 | 0 |
| y-Eudesmol | 1145 | 0 | 0 | 0 | 0.12 | 0 | 0 | 0 | 0.6 | 0 | 0.78 | 1.79 | 0 |
| tau.-Cadinol | 1026 | 0 | 0 | 0 | 0 | 0.23 | 0 | 0 | 0 | 0 | 0.25 | 1.25 | 0.37 |
| y-Cadinene | 856 | 0 | 0 | 0 | 0 | 0.56 | 0 | 0 | 0 | 0 | 0 | 0 | 0.28 |
| Isothymol methyl ether | 1234 | 0 | 0 | 0 | 0 | 1.07 | 0 | 0 | 0 | 0 | 0 | 0 | 0.96 |
| (+)-2-Carene | 1126 | 0 | 0 | 0 | 0 | 0.21 | 1.63 | 0 | 0 | 0 | 0.16 | 0 | 0.19 |
| 6-Cadinene, (+)- | 975 | 0 | 0 | 0 | 0 | 0 | 1.3 | 1.25 | 0.48 | 2.03 | 1.89 | 0.48 | 0 |
| endo-Borneol | 1596 | 0 | 0 | 0 | 0 | 0 | 0 | 3.64 | 0.41 | 2.67 | 1.96 | 0.52 | 0.97 |
| Caryophyllene oxide | 1138 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.18 | 2.65 | 1.12 | 0.32 | 0.89 |
| 84.25 | 83.58 | 88.91 | 89.21 | 83.27 | 81.09 | 86.59 | 86.12 | 81.3 | 84.65 | 90.32 | 91.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



