
Submitted:
04 July 2025
Posted:
07 July 2025
You are already at the latest version
Abstract
Diploid Vaccinium fuscatum is a wild blueberry species with low chilling requirement, evergreen growth habit, and soil adaptability to US southeast growing regions. Regardless of its potential to improve the abiotic and biotic resilience of cultivated blueberries, this species has rarely been used for blueberry breeding. One hurdle is the ploidy barrier between diploid V. fuscatum and tetraploid cultivated highbush blueberries. To overcome the ploidy barrier, vegetative shoots micro-propagated from one genotype of V. fuscatum selected because it grew vigorously in vitro and two southern highbush cultivars, ‘Emerald’ and ‘Rebel,’ were treated with colchicine. While shoot regeneration was severely repressed in ‘Emerald’ and ‘Rebel,’ shoot production from the V. fuscatum clone was not compromised at either 500 µM or 5,000 µM colchicine concentrations. Due to the high number of shoots produced in vitro by the V. fuscatum clone shoots of this clone that had enlarged stem diameter in vitro were subjected to flow cytometer analysis to screen for induced polyploidy. Sixteen synthetic tetraploid V. fuscatum, one synthetic octoploid ‘Emerald,’ and three synthetic octoploid ‘Rebel’ were identified. Growth rates of the polyploid induced mutants were reduced compared to their respective wild type controls. Leaf width and length of synthetic tetraploid V. fuscatum and synthetic octoploid ‘Emerald’ was increased compared to the wildtypes whereas the leaf width and length of synthetic octoploid ‘Rebel’ was reduced compared to the wildtype controls. Significant increases in stem thickness and stomata guard cell length were found in the polyploidy-induced mutant lines compared to the wildtypes. In the meantime, stomata density was reduced in the mutant lines. These morphological changes may improve drought tolerance and photosynthesis in these mutant lines. Synthetic tetraploid V. fuscatum can be used for interspecific hybridization with highbush blueberries to expand the genetic base of cultivated blueberries.
Keywords:
polyploidy induction
; tissue culture
; colchicine
; flow cytometry
; blueberries
1. Introduction
Blueberries have many health benefits such as improving cognitive and digestive health, and preventing cardiovascular diseases, diabetes, and muscular injuries [1]. Public awareness of these health benefits led to a steady increase in blueberry consumption and market demand [2]. As one of the world leading blueberries producers, the United States produced 333,660 ton of blueberries in 2023, which was three times more than the production in 2000 [3]. US blueberry industry generated $4.7 billion of economic impact by sustaining over 44,000 full time jobs each year [4]. It is predicted that the market demand for blueberry will continue to rise in the coming years due to the year-round market availability of blueberries, convenience for consumption, and awareness of health benefits among consumers [5]. To meet the demand of the expansion of the blueberry industry, developing high yielding blueberry cultivars not only adapted to local growing environment but also with broad harvest windows is imperative.
Blueberries (Vaccinium sect. Cyanococcus) are native to North America [6]. Major types of cultivated blueberries include northern highbush and southern highbush (both interspecific hybrids based on V. corymbosum, 2n=4x=48, NHB and SHB), lowbush (V. augustifolium, 2n=4x=48, LB), and rabbiteye blueberries (V. virgatum, 2n=6x=72, RE). V. sect. Cyanococcus was divided into 9 diploid, 12 tetraploid, and 3 hexaploid species based on morphological characteristics by Camp [6]. Vander Kloet [7] later lumped 9 highbush species at diploid, tetraploid, and hexaploid levels into V. corymbosum Efforts are underway to resolve the taxonomic dispute based on phylogenetic analysis and comprehensive sampling [8]. In this study, we adopted the Camp treatement since it is widely used in the relavent literature.
Interspecific hybridization among cultivated species and wild germplasm played a key role in blueberry cultivar development. Domestication and breeding of highbush blueberry started with interspecific hybridization between V. corymbosum and V. angustifolium in the early 1900’s [9]. Backcrosses to wild highbush selections from New Jersey and intercrosses among selected progenies resulted in the release of northern highbush blueberry cultivars [10,11]. Subsequent interspecific hybridization enabled the expansion of blueberry cultivation to a wide range of soils and climate conditions [12]. This was exemplified by the development of SHB blueberries that thrive in regions with warmer climate through introgression of low-chill and soil adaptability from V. virgatum and V. darrowii (2n=2x=24) into NHB genetic background [13]. Besides V. virgatum and V. darrowii, another diploid species V. elliottii (2n=2x=24) contributed to the fruit flavor, fragrance, upland soil tolerance and disease resistance of SHB cultivars such as ‘Snowchaser’, ‘Carteret’, and ‘Kestrel’ [14]. Signatures of introgression from both hexaploid and diploid wild blueberries were identified in SHB through population structure analysis [15].
Compared to the active breeding efforts with V. virgatum, V. darrowii, and V. elliottii, little attention has been paid to V. fuscatum, another wild blueberry species native to the southeastern U.S. Wild tetraploid populations of native V. corymbosum occur in abundance on favorable sites as far south as Gainesville in the northern part of the Florida peninsula. Native highbush blueberry plants of similar morphology, ecology, and phenology occur for another 350 km south of where the tetraploid highbush reach their southern limit. Camp [16] called the highbush blueberries from these southern locations V. fuscatum, but did not have the opportunity to determine whether they were diploid or tetraploid. Vander Kloet [17], who spent several months studying the native blueberries surrounding the Archbold Biological Station in Arcadia, Florida, determined that V. fuscatum from south Florida was all diploid. These diploid V. fuscatum plants in southwest Florida are evergreen with 1 to 5 major canes, 2 to 4 m tall, and produce small, black, sweet, juicy berries in abundance [18]. Natural habitats of V. fuscatum include sandy flatwoods, bottom lands, banks along streams or lakes [16]. Morphological characteristics of lanceolate-elliptic leaves, evergreen growth habit, and small black fruits distinguish V. fuscatum from other Vaccinium species (Figure 1). Although there is a high level of phenotypic diversity within this species, individual plants from V. fuscatum genetically formed a distinct cluster from V. elliottii and V. darrowii by principal coordinate analysis using expressed sequence tag-simple sequence repeats (EST-SSR) markers [19].
Since diploid highbush blueberry plants of similar type extend northward on the coastal plain all the way to the northeastern United States [20], it seems likely that the V. fuscatum of south Florida will eventually be found to be conspecific with the diploid highbush blueberry designated as V. atrococcum in areas north of Florida. Previously, a diploid V. fuscatum clone (reported as V. atrococcum Heller) with high level of field resistance to Phytophthora cinnamomi was identified [21]. There is a strong triploid block of producing infertile progenies from crosses between diploid and tetraploid blueberry species [22,23]. Very few viable hybrids were recovered from V. fuscatum and SHB crosses [18]. To circumvent the triploid block, axillary buds from this disease resistant clone were treated in vivo with colchicine and induced a chimeral plant with some tetraploid branches [24]. Field resistance to phytophthora root rot was retained in the colchicine-induced tetraploid clone [24]. Unfortunately, this valuable V. fuscatum clone is not available anymore.
Besides resistance to Phytophthora cinnamomi, V. fuscatum is of interest in blueberry breeding because it has the plant architecture of cultivated highbush blueberry, comes from a place which receives essentially no winter chilling, and grows as an evergreen plant [25]. Evergreen production systems for SHB blueberries are established in regions with mild winters such as California, Florida, Hawaii in the U.S., and other countries including Spain, China, Morrocco, Mexico, Argentina, and Australia [26,27,28]. Low chilling requirement, disease, and pest resistance are required to keep healthy leaves all year round for evergreen productions. Blueberry leaf rust (Thekopsora minna) is a major fungal pathogen which causes defoliation and yield reduction in the evergreen blueberry production in Florida [29] and Australia [30]. Besides blueberry rust, anthracnose, Septoria, and target spots can also cause premature leaf defoliation and yield reduction in blueberries in Florida. As a Florida native species with evergreen growth habit, V. fuscatum is tolerant or resistant to multiple blueberry foliar diseases. Utilization of V. fuscatum to improve the vigor and resistance of SHB blueberries to foliar diseases is highly desirable. In this study, we identified a robust V. fuscatum clone with evergreen growth habit and good regeneration capability in tissue culture (Figure 1). To overcome the ploidy barrier between this species and SHB, we induced polyploidy of the V. fuscatum clone through in vitro colchicine treatment.
Polyploidy induction through colchicine treatment effectively produced fertile synthetic tetraploids from diploid V. elliottii and V. darrowii as reviewed recently [31]. Colchicine is an anti-mitotic agent which disrupts spindle formation during mitosis, resulting in cells with an additional set of chromosomes [32]. Optimization of colchicine treatment conditions is necessary to reduce the lethality of treated materials from the cytotoxic effect of colchicine and improve the recovery rate of polyploidy-induced materials [33]. A wide range of colchicine concentrations from 2.5 µM to 12,518 µM were used for polyploid induction of Vaccinium species [24,34,35,36]. Regeneration of polyploidy-induced mutants from colchicine treatment was genotype specific. For instance, colchicine concentration at 2.5 µM was found to be optimum for producing synthetic octoploid mutant clones from NHB cultivar ‘Duke’ [35]; whereas 250 µM colchicine was effective in inducing synthetic tetraploids from V. elliottii and V. darrowii [37]. 500 µM and 5,000 µM colchicine concentrations were chosen for this experiment based on previously reported effective colchicine concentrations [34,36]. Increasing the ploidy level of diploid V. fuscatum to tetraploid could potentially improve the efficiency of interspecific hybridization and allele introgression. The objectives of this research are 1) evaluating the efficiency of polyploid induction of V. fuscatum in comparison with SHB cultivars; 2) recovering synthetic tetraploid V. fuscatum upon colchicine treatment; 3) characterizing morphological changes in the synthetic tetraploid V. fuscatum. The success in inducing polyploid in diploid V. fuscatum reported in this study opens an efficient venue to utilize this valuable wild blueberry species for blueberry cultivar improvement.
2. Material and Methods
2.1. Blueberry Plant Materials
In July 2017 softwood cuttings for clonal propagation were taken from 30 diploid highbush blueberry plants growing in southwest Florida. About half of the plants came from a wet forest in southwestern DeSoto County near Ft Myers, Florida. Two plants came from a wetland southeast of Frostproof, Florida and ten were selected from the wet margins of Lake Istokpoga near Sebring, Florida. The cuttings were rooted and grown in 5-gallon pots of peat. In January 2019, 30 potted plants, one from each genotype, were placed outside with several dozen honeybee hives near the University of Florida entomology department building, Gainesville, FL, where they were isolated from other diploid blueberry plants. One-hundred mature berries were harvested from each plant. The seeds were extracted in bulk and germinated in a greenhouse in Gainesville, FL. Approximately fifty of the resulting seedlings were planted at the University of Florida Plant Science Unit in Citra, Florida in May 2020. In August 2023, soft new tissue was taken from ten plants that had the largest leaves and most upright stature to establish tissue cultures in Tifton, Georgia. Tissue culture was established for six out of the ten plants. V. fuscatum clone ‘FL 21-1423’ was selected for colchicine treatment due to its high vigor in tissue culture (Figure 1). SHB blueberry cultivars ‘Emerald’ [38] and ‘Rebel’ [39] established in tissue culture by Fall Creek Nursery (Lowell, OR USA) were treated together V. fuscatum ‘FL 21-1423’ to compare the efficiency of polyploidy induction. V. fuscatum ‘FL 21-1423’ was simplified as V. fuscatum for the rest of the writings, however, additional research will be needed to determine the representativeness of this clone for V. fuscatum.
2.2. Tissue Culture and Colchicine Treatment
To avoid the high contamination rate from using plant tissue collected directly from the field [40], tissue from new axillary shoots grown in a controlled environment was used to initiate tissue culture. In this process, softwood cuttings from the V. fuscatum clone grown in the field were defoliated and washed with mild hand soap by using a paintbrush for 3 minutes. The cleaned shoots were rinsed with tap water and transferred to a beaker filled with tap water. The shoots were placed on a rack next to the window of the lab with a constant temperature of 24 ℃. Once axillary shoots grew to approximately 20 mm long in about a month, they were harvested and defoliated. The shoots were immersed in 50 ml Conical tubes with 30% Clorox and 0.005% Tween-20 and agitated for 20 minutes. The shoots were transferred to new sterile tubes and rinsed with sterilized water 3 times for 3 minutes per rinse. The shoots were segmented into single-node segments and placed horizontally in individual 15 ml tubes with 3 ml of woody plant medium (Phytotech, Lenexa, KS, USA) containing 13.7 µM zeatin (Phytotech). The WPM medium was made by adding 3% sucrose (Aldon, Avon, NY, USA) and adjusted the pH to 5.2 with NaOH. 0.8% agar (Sigma Aldrich, St. Louis, MO. USA) was added and before autoclaving at 121°C for 20 minutes. Zeatin was supplemented after the medium cooled to 60℃. To determine the lethality of colchicine treatment, V. fuscatum and SHB cultivars ‘Rebel’ and ‘Emerald,’ were treated with WPM liquid media supplemented with 0, 500 µM, and 5000 µM colchicine solution for 48 hours. During the treatment, the 50 ml tubes containing the shoots with various colchicine solutions were rocked at 30 rpm on a UltraRock rocking platform (Bio-Rad Laboratories, Hercules, CA, USA). The shoots were washed with sterile deionized water 5 times for 3 minutes per wash. Single-node stem segments were placed horizontally on the WPM medium with 2.3 µM zeatin for axillary shoot induction. For each genotype/treatment combination, there were three experimental replicates with each replicate consisting of five single-node segments arranged in one petri dish. Shoot number and shoot length were measured for each regenerated shoot 100 days after the colchicine treatment. Tissue culture plates were grown in the I-66LLVL biological incubator (Percival Scientific, Perry, Iowa, USA) at 26°C with photon flux density 40- 50 µmol/m2/s with 16/8 day and night cycles.
2.3. Rooting the Shoots from Tissue Culture
Shoots of 2 to 3 cm long were separated from the tissue culture clusters and transplanted to the 72-cell propagation tray (https://www.bootstrapfarmer.com) filled with pine bark medium. The propagation tray was enclosed with a clear cover to maintain the high humidity for four weeks in the heated greenhouse set at 18 to 28°C. Upon root formation, the plantlets were transferred to 1-gallon pots and fertilized as needed with the slow-release 10-10-10 fertilizer (Scotts Miracle-Gro Company, Marysville, OH, USA).
2.4. Flow Cytometry Analysis
Two to three leaves per shoot regenerated in tissue culture were dissected for ploidy level determination. The protocol from CyStain® PI Absolute P staining kit (Sysmex Partec GmbH, Görlitz, Germany) was modified to adapt to the 5-10 mg of tissue harvested from shoot explants. Equal amount of leaf tissue from target plantlet and control were chopped together in 60 µl of extraction buffer for 30 to 60 seconds in a Petri dish chilled on ice. 500 µl of working staining solution (0.5 µl staining buffer, 3 µl propidium iodide (PI) dye, 1.5 µl of RNase and 495 µl of water) was added to the nuclei extract and incubated for 60 seconds. The PI-labeled nuclei solution was filtered through a 30 µm CellTrics filter (Sysmex Partec GmbH) and incubated on ice for 10 minutes before running through the Attune NxT Acoustic Focusing Cytometer (ThermoFisher, Scientific, Walthman, MA, USA). X-mean confidence interval less than 5% were included in ploidy determination using the following formula. X-mean value is the mean propidium iodide (PI) fluorescence signal emitted by the PI-labeled nuclei. This value indicates the intensity of the fluorescent signal of the target nuclei. To determine the ploidy level of colchicine treated V. fuscatum, tissue from RE was used as a control (Figure 2A and B). To determine the ploidy level of colchicine treated SHB, diploid V. fuscatum was used as the control.
Ploidy level of solid synthetic polyploids was confirmed by additional flowcytometry analysis with the new shoots micro-propagated from the putative shoots.
2.5. Morphological Characterization
Axillary shoots collected from tissue culture were imaged in a sterile laminar flow hood by a digital microscope (Moysuwe, https://www.amazon.com/MOYSUWE-Microscope-Magnifier-Soldering-Compatible/dp/B0CB2J33SB?ref_=ast_sto_dp) before leaf sample collection for flow cytometry analysis. Stem thickness of these shoots was measured by image analysis using APS Assess 2.0 (The American Phytopathological Society, St. Paul, MN, USA). After the tissue cultured shoots were well-established in the greenhouse, leaf samples were collected from seven-month-old plants for stomata, guard cell and leaf size measurements. For stomata impressions, the third leaf from the top was collected. A thin layer of clear nail polish was applied to the abaxial side of the leaf. Once dried, the stomata impression was lifted from the leaf tissue by applying a clear tape. The impression was subsequently loaded onto a microscope slide and viewed under a Nikon Eclipse Si microscope (Nikon Corporation, Tokyo, Japan). Three fields of view were recorded for stomata density from each leaf. Three leaves per genotype were recorded. The length and width of the guard cells were recorded from nine stomates with closing pores from each genotype using APS Assess 2.0. For leaf size, images of another twelve mature leaves were imported into the APS Assess 2.0 software to determine leaf width and length. For plant height, the length of the main cane was measured to represent plant height. The number of canes produced by each plant was counted.
2.5. Statistical Analysis
To determine the effect of polyploidy induction on plant morphology, a student t-test was performed to compare the measurements between the synthetic polyploids and their respective wild type plants. Statistical significance was determined at P<0.05. Normality tests were performed using the Shapiro–Wilk test, and the datasets were found to be normally distributed.
3. Results and Discussion
For the colchicine lethality experiment, diverse genotypic responses to axillary shoot induction was observed the WPM medium without colchicine, which was consistent with the previous report on Vaccinium species [41]. ‘Rebel’ produced callus at the base of the new shoots whereas callus formation was minimum for ‘Emerald’ and V. fuscatum (Figure 3). Callus formation at the base of the axillary shoot is undesirable in this case since none of the callus tissue further differentiated and produced adventitious shoots [42]. V. fuscatum exhibited vigorous growth and produced an average of eight shoots per nodal segment which was significantly higher than ‘Rebel’ and ‘Emerald’ whose average shoots per segment was four (Figure 4A). No significant difference for axillary shoot length was found among these genotypes without colchicine treatment (Figure 4B). This result confirmed our initial observation of the vigorous shoot growth of this V. fuscatum clone while establishing tissue culture from softwood cuttings (Figure 1D). When the nodal segments were treated with 500 µM and 5000 µM of colchicine, significant suppression of shoot induction and elongation was found in all three genotypes (Figure 3). At 500 µM colchicine level, V. fuscatum produced an average of 4.7 shoots per segment which was significantly higher than ‘Rebel’ and ‘Emerald’ whose average shoot number per segment was reduced to 0.6 and 0.3 respectively (Figure 4A). In addition, the shoot length of V. fuscatum was also significantly longer than ‘Rebel’ and ‘Emerald’ (Figure 4B). At 5000 µM colchicine level, the average number of shoots per nodal segment was reduced to 0.2, 0.3, and 0.9 for V. fuscatum, ‘Rebel,’ and ‘Emerald’ which was no significantly different across the genotypes. There was no significant genotypic difference on shoot length at 5000 µM colchicine level and the average shoot length was reduced to less than 6 mm. In this experiment, although suppressive effect of colchicine on plant growth was found at both 500 and 5000 µM levels, V.fuscatum survived better than SHB cultivars at 500 µM colchicine level by producing more vigorous axillary shoots.
Colchicine is a natural tri-cyclic alkaloid extracted from Gloriosa superba and Colchicum autumnale in the Liliaceae family [43]. When actively dividing plant cells are treated with colchicine, cell division is arrested due to the binding of colchicine with tubulins which interrupts microtubules formation during the metaphase of mitotic cell cycle [44]. The absence of chromosomal pair separation results in the retention of two sets of chromosomes in the mutant cell, i.e., polyploidy induction [45]. Lethality of colchicine depends on the concentration and duration of colchicine treatment [46]. In our experiment, the suppressive effect of colchicine on shoot induction cognates with the previous findings [37,47]. The higher survival rate of V. fuscatum suggested that it has a higher tolerance to the mitotic agent compared to the SHB cultivars.
Due to the low shoot induction rate upon colchicine treatment, the number of nodal segments for colchicine treatment was increased to ensure the recovery of induced polyploids (Table 1).
Seventy-two nodal segments of ‘Rebel’ and ‘Emerald’ were treated with 500 µM colchicine whereas 200 nodal segments each were treated with 5000 µM colchicine. As for V. fuscatum, seventy-nine nodal segments of were treated for both colchicine levels. Shoots regenerated from colchicine treated nodal segments of both ‘Rebel’ and ‘Emerald’ grew slowly. It took three transfers in four months to establish 97 and 116 shoots for ‘Rebel’ and ‘Emerald’ respectively. The average axillary shoots per treated segment ranged from 0.3 to 0.6 for these two cultivars which was similar to the results of the colchicine lethality experiment (Figure 3). Ploidy levels of all the regenerated shoots from these cultivars were determined by flowcytometry analysis. Three synthetic octoploid ‘Rebel’ were identified among the thirty regenerated shoots from the 500 µM colchicine treatment. No synthetic octoploid was recovered from the sixty-seven ‘Rebel’ shoots regenerated from the 5000 µM colchicine treatment. As for ‘Emerald,’ only one octoploid was identified among the seventy-six regenerated shoots from 5000 µM colchicine treatment and there was no synthetic octoploid among the forty shoots regenerated from 500 µM colchicine treatment. The overall success rate of polyploid induction was 3% and 0.9% for ‘Rebel’ and ‘Emerald’ respectively. These success rates of polyploid induction were similar to the reported rates of other SHB cultivars, ‘Legacy,’ ‘Duke’, and ‘Biloxi’ [48]. Besides octoploids, 9 and 4 mixoploids were identified in ‘Rebel’ and ‘Emerald’ respectively. Blueberry mixoploids are often discarded due to their genome instability [49]. Since the number of synthetic octoploids was low, we further recovered more synthetic octoploids from these mixoploids through chimera dissociation by leaf organogenesis and axillary shoot induction in a separate study [50].
As for V. fuscatum, when the number of treated nodal segments increased to seventy-nine, it exhibited a highly robust growth and completely overcome the suppressive effect from both levels of colchicine treatments. Nine-weeks after the colchicine treatment, V. fuscatum produced 676 and 632 new shoots at 500 µM and 5000 µM colchicine levels respectively, which was an average of 8.6 and 8 shoots per segment. This level of shoot induction was comparable to the level of shoot induction without colchicine treatment (Figure 3). The exuberant growth of colchicine-treated V. fucatum upon sample size increase indicated that V. fuscatum has an exceptional resilience to detoxify the cytotoxicity of colchicine. Previously, transcriptome analysis of colchicine treated plants revealed that besides upregulating of genes inhibiting the formation of microtubule, spindle, and chromosomal kinetochore, colchicine also decreased cytokinesis resulting in reduced cell activity and promoting apoptosis [51]. In addition to the application in plant biology, colchicine has been used to treat inflammatory conditions such as gout disease clinically [52]. Overdose or misuse often resulted in colchicine poisoning among patients [53]. The mechanism of detoxification against colchicine treatment in V. fuscatum needs to be studied further since it may provide an alternative therapeutic option for colchicine poisoning.
Regardless, this large amount of shoot induction in V. fuscatum made it challenging to screen all of them for flowcytometry analysis. Previously, enlargement of stem diameter was reported for polyploid induced Vaccinium species such as V. elliottii [54], V. darrowii [37], highbush blueberries [33], and V. virgatum [36]. Consequently, 35 and 33 shoots with thicker stems from 500 µM and 5000 µM colchicine treatments respectively were visually selected for flowcytometry analysis (Table 1). A total of 16 synthetic tetraploid shoots were identified accounting for 24% of the evaluated shoots. Therefore, the preferential selection of V. fuscatum shoots with thick stem resulted in a higher recovery rate of synthetic polyploids than the success rates from ‘Rebel’ and ‘Emerald’ in our study. This success rate was also higher than the highest success rate of 11% reported in literature for wild blueberry species V. myrtillus [55] and V. corymbosum [56]. Therefore, in the case of genotypes highly prolific in shoot induction upon colchicine treatment, it is advisable to speed up the identification of polyploidy-induced mutant lines by visually selecting shoots with thicker stems for flowcytometry analysis.
Producing multiple shoots carrying the same mutation is desirable due to the potential loss of the valuable mutant line to the subsequent micropropagation, rooting, and cultivation processes. These synthetic polyploids are genetically stable and ready for breeding or clonal propagation. However, from the standpoint of creating a diverse genetic resource of V. fuscatum at tetraploid level, it may be more efficient to treat the seed population instead of micro-propagated individual clones and treatment them with colchicine. Previously, 0.04% to 5% recovery rates were reported when seeds from Vaccinium species were treated with colchicine [47,57,58,59,60,61]. Due to the genetic heterozygosity within Vaccinium species [22], the polyploidy-induced mutant lines from individual seeds would carry unique genomic compositions. However, the downside of this alternative method lies in the challenge of differentiating solid polyploidy induced mutant lines from sectorial, periclinal, and mericlinal chimeras [58].
Polyploidy induction was reported to increase the size and weight of multiple plant organs such as stem, fruit, flower, and leaves [62]. This type of phenotypic shift has been accredited to the allele dosage effect in the mutant lines [63]. Since only four octoploid shoots were recovered from ‘Rebel’ and ‘Emerald’, stem thickness from these two SHB cultivars were combined to determine the effect of polyploidy. Synthetic octoploid SHB had significantly thicker stems than the wildtype tetraploid SHB (Figure 5A). Similarly, stems of synthetic tetraploid V. fuscatum were significantly thicker compared to the wildtype diploid V. fuscatum (Figure 5B). These results are consistent with the previous findings of enlarged stem diameter from colchicine treatment [64,65,66]. Mixoploids were recovered from both SHB and V. fuscatum. The stem thickness of the mixoploids was numerically in between the polyploid-induced mutant and wildtype controls for both species (Figure 5). For the SHB samples, the thickness of mixoploid stems was statistically significantly lower than the synthetic octoploid but similar to the wildtype control. In the case of V. fuscatum, the stem thickness of mixoploid was not statistically different from either the synthetic tetraploid V. fuscatum or the wildtype control.
The in vitro micro-propagated shoots were rooted and established in pine bark medium in the greenhouse (Figure 6). Plant morphological characteristics were measured for these plants (Table 2). Upon polyploid induction, both plant height and the number of canes were reduced in all three polyploidy-induced genotypes compared to their respective wildtype controls (Table 2). The differences reached statistical significance except for the number of canes for synthetic tetraploid V. fuscatum. These data indicate that the polyploid induction reduced the plant growth of the genotypes included in this study. Similarly, an increase in polyploid level was reported to reduce the growth rate of Arabidopsis [67]. As for leaf size, both synthetic tetraploid V. fuscatum and octoploid ‘Emerald’ had signficantly increased leaf width and length compared to their respectively wild type controls; except for the increment in leaf length of octoploid ‘Emerald’ did not reach statistical significance. On the contrary, both leaf width and length of synthetic octoploid ‘Rebel’ were significantly smaller than the wildtype control. The increment of leaf length and withdth in the polyploidy-induced mutants was reported to associate with increase in cell size and cell elongation contributed by the ‘gigas effect” from genome doubling [68,69]. In our study, the opposite effect of polyploidy induction on the leaf width and length in ‘Rebel’ could be specific to the polyploidization events created in this study or it could be genotype-specific response to the polypoid induction. Diverse genotypic response to polyploidy induction was reported in grasses [70]. To further understand the nature of this negative response, more independent polyploidization events need to be created from ‘Rebel’ in the future. For stomata guard cell size and density, enlargement of the stomata guard cell size and reduction in stomata density was observed for both polyploidy-induced SHB and V. fuscatum (Table 2, Figure 7) Synthetic octoploid ‘Emerald’ and ‘Rebel’ had significantly wider and longer stomata guard cells compared to their respective wildtype controls. Synthetic tetraploid V. fuscatum had significantly longer stomata guard cells than the diploid wildtype whereas the numerical increment in guard cell width in the synthetic tetraploids did not reach statistical significance. Polyploidy-induced V. darrowii and V. elliottii also reported to have longer guard cells [37,58]. Statistically significant reduced stomata density was found in polyploidy-induced lines of both V. fuscatum and ‘Emerald’ (Table 2 and Figure 7) whereas the reduction of stomata density in ‘Rebel’ did not reach statistical significance. Previously, larger guard cell size was found to moderately increase water use efficiency and drought tolerance in Z. Mays [71]. Reduction in stomata density was reported in other polyploidy-induced highbush blueberries [56]. Stomata are small pores bound by a pair of guard cells on the leaf surface. They regulate water and CO2 exchange in response to the environmental cues [72]. Reduction in stomata density was found to associated with increased drought tolerance in wheat [73,74] and photosynthesis in rice [75]. As the frequency and severity of drought events are intensified due to the global climate change [76], testing the drought tolerance and photosynthetic efficiency of the synthetic polyploids in the future will inform the utility of these synthetic mutants in improving drought tolerance in blueberries.
Conclusion
Polyploidy induction through colchicine treatment successfully induced tetraploid V. fuscatum from a wild diploid clone. The high vigor of shoot induction of V. fuscatum upon colchicine treatment not only enabled highly efficient screening of polyploidy-induced mutant lines but also could be useful for studying the mechanism of detoxifying the cytotoxicity of colchicine. The synthetic tetraploid V. fuscatum had morphological changes including increased stem thickness and stomata guard cell size, and reduced stomata density. These characteristics may increase drought tolerance and photosynthetic efficiency in the synthetic tetraploid V. fuscatum. From the perspective of breeding, it is expected that the synthetic tetraploid V. fuscatum will produce viable pollen and egg cells that can be used for interspecific hybridization with highbush blueberries. This effort will contribute to the expansion of genetic diversity of cultivated blueberries to meet the challenges of blueberry production.
Funding
The authors would like to acknowledge the USDA-NIFA Hatch project (project number: 7004954) for funding the study.
Acknowledgments
The authors want to thank Tracey Cook and Sindoora Nalajala for their technical assistance in data collection.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Stull, A.J.; Cassidy, A.; Djousse, L.; Johnson, S.A.; Krikorian, R.; Lampe, J.W.; Mukamal, K.J.; Nieman, D.C.; Porter Starr, K.N.; Rasmussen, H. The state of the science on the health benefits of blueberries: a perspective. Frontiers in Nutrition 2024, 11, 1415737. [Google Scholar] [CrossRef] [PubMed]
- Hammami, A.M.; Guan, Z.; Cui, X. Foreign competition reshaping the landscape of the US blueberry market. Choices 2024, 39, 1–9. [Google Scholar] [CrossRef]
- FAOSTAT. 2023. doi:https://www.fao.org/faostat/en/#data/QCL/visualize.
- Tootelian, D. National and state-specific economic impact study. 2023, https://ushbc.blueberry.org/allresources, doi:https://ushbc.blueberry.org/all-resources.
- IBO. Global Fresh Blueberry Outlook 2025–2030. 2025. doi:https://www.internationalblueberry.org/2025/04/14/global-fresh-blueberry-outlook-2025-2030/.
- Camp, W.H. The North American blueberries with notes on other groups of Vacciniaceae. Brittonia 1945, 5, 203–275. [Google Scholar] [CrossRef]
- Vander Kloet, S.P. The genus Vaccinium in north America; Ottawa, ON, Canada, 1988.
- Fritsch, P.W.; Crowl, A.A.; Ashrafi, H.; Manos, P.S. Systematics and evolution of Vaccinium Sect. cyanococcus (Ericaceae): progress and prospects. Rhodora 2024, 124, 301-332, 332. [CrossRef]
- Moore, J.N. Improving highbush blueberries by breeding and selection. Euphytica 1965, 14, 39–48. [Google Scholar] [CrossRef]
- Coville, F.V. Blueberry chromosomes. Blueberry chromosomes. Science 1927, 66, 565-566. [CrossRef]
- Longley, A. Chromosomes in Vaccinium. Science 1927, 66, 566–568. [Google Scholar] [CrossRef]
- Draper, A. Blueberry breeding: improving the unwild blueberry. J. Am. Pom. Soc. 2007, 61, 140–143. [Google Scholar]
- Lyrene, P.M. Value of various taxa in breeding tetraploid blueberries in Florida. Euphytica 1997, 94, 15–22. [Google Scholar] [CrossRef]
- Norden, E.H.; Lyrene, P.M.; Chaparro, J.X. Ploidy, fertility, and phenotypes of F1 hybrids between tetraploid highbush blueberry cultivars and diploid Vaccinium elliottii. HortSci. 2020, 55, 281–286. [Google Scholar] [CrossRef]
- Nishiyama, S.; Fujikawa, M.; Yamane, H.; Shirasawa, K.; Babiker, E.; Tao, R. Genomic insight into the developmental history of southern highbush blueberry populations. Heredity 2021, 126, 194–205. [Google Scholar] [CrossRef]
- Camp, W.H. Description of species: Vaccinium darrowi-Vaccinium hirsutum. Brittonia 1945, 5, 220–266. [Google Scholar] [CrossRef]
- Vander Kloet, S.P. The taxononmy of Vaccinium and cyancoccus: a summation. Can. J. Bot. 1983, 61, 256–266. [Google Scholar] [CrossRef]
- Lyrene, P.M. Florida native blueberries and their use in breeding. In Proceedings of the XI International Vaccinium Symposium 1180, 2016; pp. 9-16.
- Bassil, N.; Bidani, A.; Hummer, K.; Rowland, L.J.; Olmstead, J.; Lyrene, P.; Richards, C. Assessing genetic diversity of wild southeastern North American Vaccinium species using microsatellite markers. Genetic Resources and Crop Evolution 2018, 65, 939–950. [Google Scholar] [CrossRef]
- Ward, D.B. Contributions to the flora of Florida: 6, Vaccinium (Ericaceae). Castanea 1974, 191-205, doi:https://www.jstor.org/stable/4032784.
- Draper, A.; Mircetich, S.M.; Scott, D.H. Vaccinium clones resistant to Phytophthora cinnamomi. HortSci. 1971, 6, 167–169. [Google Scholar] [CrossRef]
- Ballington, J.R. The role of interspecific hybridization in blueberry improvement. In Proceedings of the IX International Vaccinium Symposium 810, 2008; pp. 49-60.
- Lyrene, P.M.; Vorsa, N.; Ballington, J.R. Polyploidy and sexual polyploidization in the genus Vaccinium. Euphytica 2003, 133, 27–36. [Google Scholar] [CrossRef]
- Draper, A.; Stretch, A.W.; Scott, D.H. Two tetraploid sources of resistance for breeding blueberries resistant to phytophthora cinnamomi Rands. HortSci. 1972, 7, 266–268. [Google Scholar] [CrossRef]
- Lyrene, P.M.; Sherman, W.B. Horticultural characteristics of native Vaccinium darrowii, V. elliottii, V. fuscatum, and V. myrsinites in Alachua County, Florida. J. Am. Soc. Hort. Sci. 1980, 105, 393–396. [Google Scholar] [CrossRef]
- Wright, G. Performance of southern highbush and rabbiteye blueberries on the Corindi Plateau NSW Australia. In Proceedings of the V International Symposium on Vaccinium Culture 346, 1993; pp. 141-146.
- Hummer, K.; Zee, F.; Strauss, A.; Keith, L.; Nishijima, W. Evergreen production of southern highbush blueberries in Hawai'i. Journal of the American Pomological Society 2007, 61, 188, doi:https://www.proquest.com/scholarly-journals/evergreen-production-southernhighbush/docview/209773128/se-2?accountid=14537.
- Brazelton, C. World blueberry acreage & production. Folsom: USHBC 2013, 353, 880-886, doi:https://pdfs.semanticscholar.org/aa0c/b8a984772846b938dbebf85b97d36e6b2afe.pdf.
- Harmon, P.F.; Liburd, O.E.; Dittmar, P.; Williamson, J.G.; Phillips, D. 2024 Florida blueberry integrated pest management guide, HS1156. UF/IFAS Extension 2024, 2024. [Google Scholar] [CrossRef]
- Simpson, M.; Wilk, P.; Collins, D.; Robertson, D.; Daniel, R. Managing blueberry rust under an evergreen system. In Proceedings of the XI International Vaccinium Symposium 1180, 2016; pp. 105-110.
- Chu, Y.; Lyrene, P.M. Artificial induction of polyploidy in blueberry breeding: A review. HortScience 2025, 60, 100–110. [Google Scholar] [CrossRef]
- Blakeslee, A.F.; Avery, A.G. Methods of inducing doubling of chromosomes in plants. By treatment with colchicine. Journal of Heredity 1937, 28, 393–412. [Google Scholar] [CrossRef]
- Goldy, R.G.; Lyrene, P.M. In vitro colchicine treatment of 4x blueberries, Vaccinium sp. J. Am. Soc. Hort. Sci. 1984, 109, 336–338. [Google Scholar] [CrossRef]
- Dweikat, I.; Lyrene, P. Production and evaluation of a synthetic hexaploid in blueberry. TAG 1989, 77, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Hernández, R.; López, A.; Valenzuela, B.; D’Afonseca, V.; Gomez, A.; Arencibia, A.D. Organogenesis of plant tissues in colchicine allows selecting in field trial blueberry (Vaccinium spp. cv Duke) clones with commercial potential. Horticulturae 2024, 10, 283. [Google Scholar] [CrossRef]
- Lyrene, P.M.; Perry, J.L. Production and selection of blueberry polyploids in vitro. J. Heredity 1982, 73, 377–378. [Google Scholar] [CrossRef]
- Perry, J.; Lyrene, P. In vitro induction of tetraploidy in Vaccinium darrowi, V. elliottii, and V. darrowi x V. elliottii with colchicine treatment. J. Am. Soc. Hort. Sci. 1984, 109, 4–6. [Google Scholar] [CrossRef]
- Lyrene, P.M. ‘Emerald’ southern highbush blueberry. HortSci. 2008, 43, 1606–1607. [Google Scholar] [CrossRef]
- NeSmith, D.S. ‘Rebel’ southern highbush blueberry. HortSci. 2008, 43, 1592–1593. [Google Scholar] [CrossRef]
- Cappai, F.; Garcia, A.; Cullen, R.; Davis, M.; Munoz, P.R. Advancements in low-chill blueberry Vaccinium corymbosum L. tissue culture practices. Plants 2020, 9, 1624. [Google Scholar] [CrossRef]
- Debnath, S.C. In vitro culture of lowbush blueberry (Vaccinium angustifolium Ait.). Small Fruits Rev. 2004, 3, 393–408. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Q.; Wang, Z.; Zhang, Y. High efficiency regeneration system from blueberry leaves and stems. Life 2023, 13, 242. [Google Scholar] [CrossRef]
- Ade, R.; RAI, M.K. Colchicine, current advances and future prospects. Nusantara Bioscience 2010, 2. [Google Scholar] [CrossRef]
- Malawista, S.E. Colchicine: a common mechanism for its anti-inflammatory and anti-mitotic effects. Arthritis & Rheumatism 1968, 11, 191–197. [Google Scholar] [CrossRef]
- Salma, U.; Kundu, S.; Mandal, N. Artificial polyploidy in medicinal plants: advancement in the last two decades and impending prospects. J. Crop. Sci. Biotech. 2017, 20, 9–19. [Google Scholar] [CrossRef]
- Zhou, H.-w.; Zeng, W.-d.; Yan, H.-b. In vitro induction of tetraploids in cassava variety ‘Xinxuan 048’using colchicine. PCTOC 2017, 128, 723–729. [Google Scholar] [CrossRef]
- Haring, R.; Lyrene, P. Detection of colchicine induced tetraploids of Vaccinium arboreum with flow cytometry. In Proceedings of the IX International Vaccinium Symposium 810, 2008; pp. 133-138.
- Jarpa-Tauler, G.; Martínez-Barradas, V.; Romero-Romero, J.L.; Arce-Johnson, P. Autopolyploidization and in vitro regeneration of three highbush blueberry (Vaccinium corymbosum L.) cultivars from leaves and microstems. Plant Cell, Tissue and Organ Culture (PCTOC) 2024, 158, 18. [Google Scholar] [CrossRef]
- Miyashita, C.; Ishikawa, S.; Mii, M. In vitro induction of the amphiploid in interspecific hybrid of blueberry (Vaccinium corymbosum× Vaccinium ashei) with colchicine treatment. Sci. Hort. 2009, 122, 375–379. [Google Scholar] [CrossRef]
- Walter, E.; Biswal, A.; Ozias-Akins, P.; Chu, Y. Leaf organogenesis improves recovery of solid polyploid shoots from chimeric southern highbush blueberry. BioTech 2025, 14, 48. [Google Scholar] [CrossRef]
- Zhou, K.; Fleet, P.; Nevo, E.; Zhang, X.; Sun, G. Transcriptome analysis reveals plant response to colchicine treatment during on chromosome doubling. Scientific Reports 2017, 7, 8503. [Google Scholar] [CrossRef]
- Liantinioti, G.; Argyris, A.A.; Protogerou, A.D.; Vlachoyiannopoulos, P. The Role of colchicine in the treatment of autoinflammatory diseases. Curr Pharm Des 2018, 24, 690–694. [Google Scholar] [CrossRef]
- Deng, H.; Xiang, P.; Zhang, S.; Wu, H.; Liu, W.; Yan, H. Delayed elimination in humans after ingestion of colchicine: Two fatal cases of colchicine poisoning. Journal of Forensic Sciences 2023, 68, 1425–1430. [Google Scholar] [CrossRef]
- Dweikat, I.M.; Lyrene, P.M. Induced tetraploidy in a Vaccinium elliottii facilitates crossing with cultivated highbush blueberry. J. Am. Soc. Hort. Sci. 1991, 116, 1063–1066. [Google Scholar] [CrossRef]
- Podwyszynska, M.; Mynett, K.; Markiewicz, M.; Pluta, S.; Marasek-Ciolakowska, A. Chromosome doubling in genetically diverse bilberry (Vaccinium myrtillus L.) accessions and evaluation of tetraploids in terms of phenotype and ability to cross with highbush blueberry (V. corymbosum L.). Agron. 2021, 11, 2584. [Google Scholar] [CrossRef]
- Marangelli, F.; Pavese, V.; Vaia, G.; Lupo, M.; Bashir, M.A.; Cristofori, V.; Silvestri, C. In vitro polyploid induction of highbush blueberry through de novo shoot organogenesis. Plants 2022, 11, 2349. [Google Scholar] [CrossRef]
- Aalders, L.; Hall, I. Note on aeration of colchicine solution in the treatment of germinating blueberry seeds to induce polyploidy. Can. J. Plant Sci. 1963, 43, 107–107. [Google Scholar] [CrossRef]
- Chavez, D.J.; Lyrene, P.M. Production and identification of colchicine-derived tetraploid Vaccinium darrowii and its use in breeding. J. Am. Soc. Hort. Sci. 2009, 134, 356–363. [Google Scholar] [CrossRef]
- Lyrene, P.M. First report of Vaccinium arboreum hybrids with cultivated highbush blueberry. HortSci. 2011, 46, 563–566. [Google Scholar] [CrossRef]
- Lyrene, P.M. Phenotype and fertility of intersectional hybrids between tetraploid highbush blueberry and colchicine-treated Vaccinium stamineum. HortSci. 2016, 51, 15–22. [Google Scholar] [CrossRef]
- Tsuda, H.; Kunitake, H.; Yamasaki, M.; Komatsu, H.; Yoshioka, K. Production of intersectional hybrids between colchicine-induced tetraploid shashanbo (Vaccinium bracteatum) and highbush blueberry ‘Spartan’. J. Am. Soc. Hort. Sci. 2013, 138, 317–324. [Google Scholar] [CrossRef]
- Eng, W.-H.; Ho, W.-S. Polyploidization using colchicine in horticultural plants: A review. Sci. Hort. 2019, 246, 604–617. [Google Scholar]
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Martin, E.; Regalado, J.; Raghavan, L.; Encina, C. In vitro induction of autooctoploid asparagus genotypes. Plant Cell, Tissue and Organ Culture (PCTOC) 2015, 121, 249–254. [Google Scholar] [CrossRef]
- Widoretno, W. In vitro induction and characterization of tetraploid Patchouli (Pogostemon cablin Benth.) plant. Plant Cell, Tissue and Organ Culture (PCTOC) 2016, 125, 261–267. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, J. Colchicine-induced tetraploidy in Dendrobium cariniferum and its effect on plantlet morphology, anatomy and genome size. Plant Cell, Tissue and Organ Culture (PCTOC) 2021, 144, 409–420. [Google Scholar] [CrossRef]
- Corneillie, S.; De Storme, N.; Van Acker, R.; Fangel, J.U.; De Bruyne, M.; De Rycke, R.; Geelen, D.; Willats, W.G.T.; Vanholme, B.; Boerjan, W. Polyploidy affects plant growth and alters cell wall composition Plant Physiology 2018, 179, 74-87. [CrossRef]
- Sugiyama, S.-I. Polyploidy and cellular mechanisms changing leaf size: comparison of diploid and autotetraploid populations in two species of Lolium. Annals of Botany 2005, 96, 931–938. [Google Scholar] [CrossRef]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef]
- Głowacka, K.; Jeżowski, S.; Kaczmarek, Z. In vitro induction of polyploidy by colchicine treatment of shoots and preliminary characterisation of induced polyploids in two Miscanthus species. Industrial Crops and Products 2010, 32, 88–96. [Google Scholar] [CrossRef]
- Rui, M.; Chen, R.; Jing, Y.; Wu, F.; Chen, Z.-H.; Tissue, D.; Jiang, H.; Wang, Y. Guard cell and subsidiary cell sizes are key determinants for stomatal kinetics and drought adaptation in cereal crops. New Phytologist 2024, 242, 2479–2494. [Google Scholar] [CrossRef]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.-C.; Li, F.-M.; Zhang, T. Performance of wheat crops with different chromosome ploidy: root-sourced signals, drought tolerance, and yield performance. Planta 2006, 224, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Xiang, C.-B. Stomatal density and bio-water saving. Journal of Integrative Plant Biology 2007, 49, 1435–1444. [Google Scholar] [CrossRef]
- Fu, Y.; Yan, H.; Li, L.; Yu, Y.; Si, H.; Hu, G.; Xiao, H.; Sun, Z. Photosynthesis-related characteristics of different ploidy rice plants. Zhongguo Shuidao Kexue 1999, 13, 157-160, doi:http://www.ricesci.cn/EN/Y1999/V13/I3/157.
- Leng, G.; Hall, J. Crop yield sensitivity of global major agricultural countries to droughts and the projected changes in the future. Science of the Total Environment 2019, 654, 811–821. [Google Scholar] [CrossRef]
Figure 1.
V. fuscatum bushes grown at the University of Florida Plant Science Unit in Citra, Florida (A); a V. fuscatum branch loaded with fruits (B); ripe fruits from V. fuscatum in the top cluster compared to fruits from SHB at the bottom cluster (C); vigorous growth of the V. fustcatum clone in tissue culture medium containing 2.3 µM zeatin (D).
Figure 1.
V. fuscatum bushes grown at the University of Florida Plant Science Unit in Citra, Florida (A); a V. fuscatum branch loaded with fruits (B); ripe fruits from V. fuscatum in the top cluster compared to fruits from SHB at the bottom cluster (C); vigorous growth of the V. fustcatum clone in tissue culture medium containing 2.3 µM zeatin (D).

Figure 2.
Flowcytometry plots differentiating polyploidy-induced mutant lines from the wildtype controls. (A)Nuclei from the wild type V. fuscatum produced a 2x peak and RE control produced the 6x peak; (B) Synthetic tetraploid V. fuscatum produced a 4x peak closely adjacent to the 6x peak of the RE control; (C) Synthetic octoploid ‘Emerald’ produced an 8x peak at the far right of 2x peak produced by the wildtype V. fuscatum control.
Figure 2.
Flowcytometry plots differentiating polyploidy-induced mutant lines from the wildtype controls. (A)Nuclei from the wild type V. fuscatum produced a 2x peak and RE control produced the 6x peak; (B) Synthetic tetraploid V. fuscatum produced a 4x peak closely adjacent to the 6x peak of the RE control; (C) Synthetic octoploid ‘Emerald’ produced an 8x peak at the far right of 2x peak produced by the wildtype V. fuscatum control.
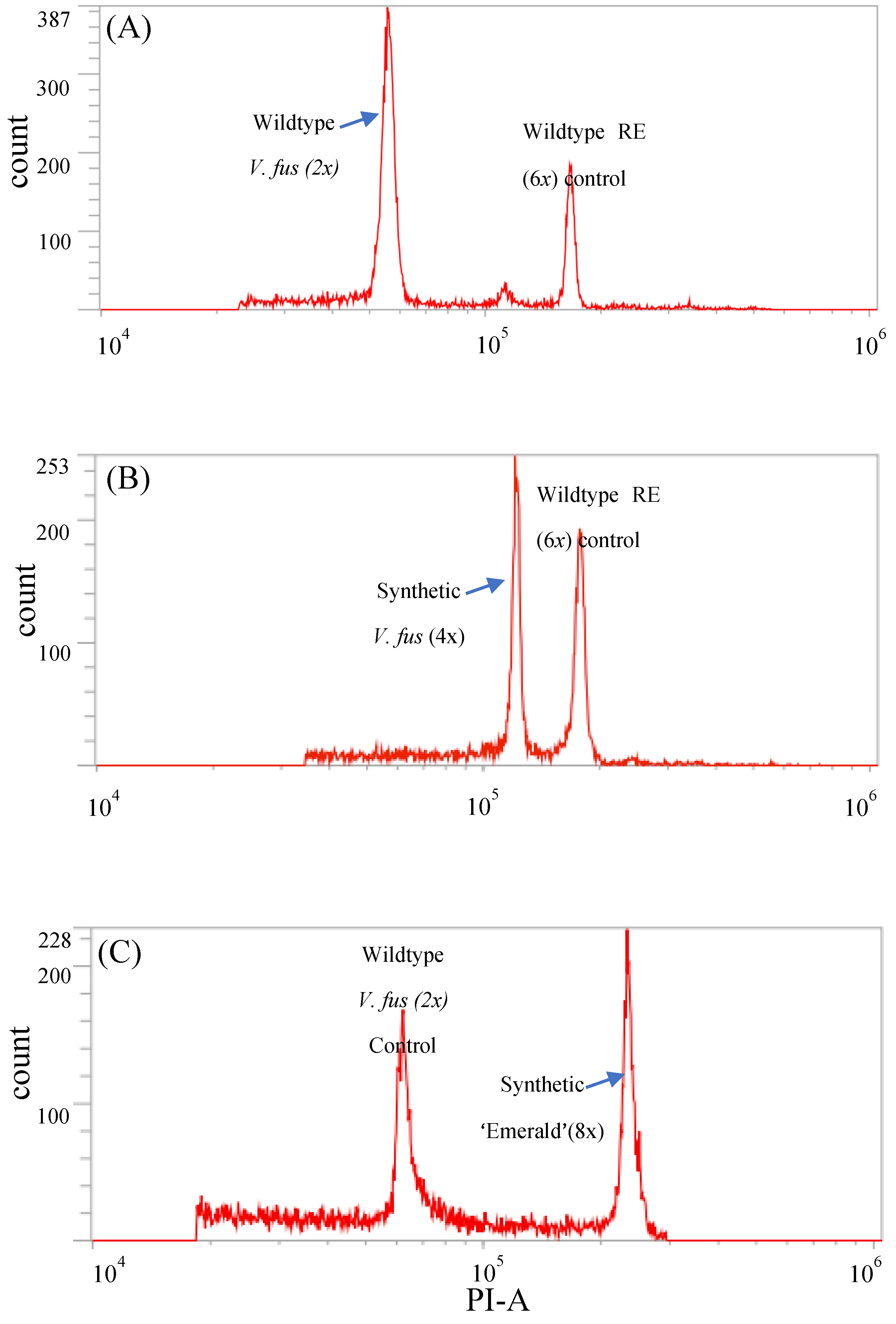
Figure 3.
Plantlets from ‘Rebel’, ‘Emerald’, and V. fuscatum micro-propagated on WPM medium not treated with colchicine (left panel) and treated with 500 µM colchicine (right panel). Images of plantlets treated with 5000 µM colchicine was omitted due to the minimum growth of all tested genotypes. Images taken at 100 days after treatment.
Figure 3.
Plantlets from ‘Rebel’, ‘Emerald’, and V. fuscatum micro-propagated on WPM medium not treated with colchicine (left panel) and treated with 500 µM colchicine (right panel). Images of plantlets treated with 5000 µM colchicine was omitted due to the minimum growth of all tested genotypes. Images taken at 100 days after treatment.

Figure 4.
Shoot number (A) and length (B) measured from axillary shoots induced from colchicine-treated nodal segments. Different letters on top of the bars indicate statistically significant difference at p<0.05.
Figure 4.
Shoot number (A) and length (B) measured from axillary shoots induced from colchicine-treated nodal segments. Different letters on top of the bars indicate statistically significant difference at p<0.05.
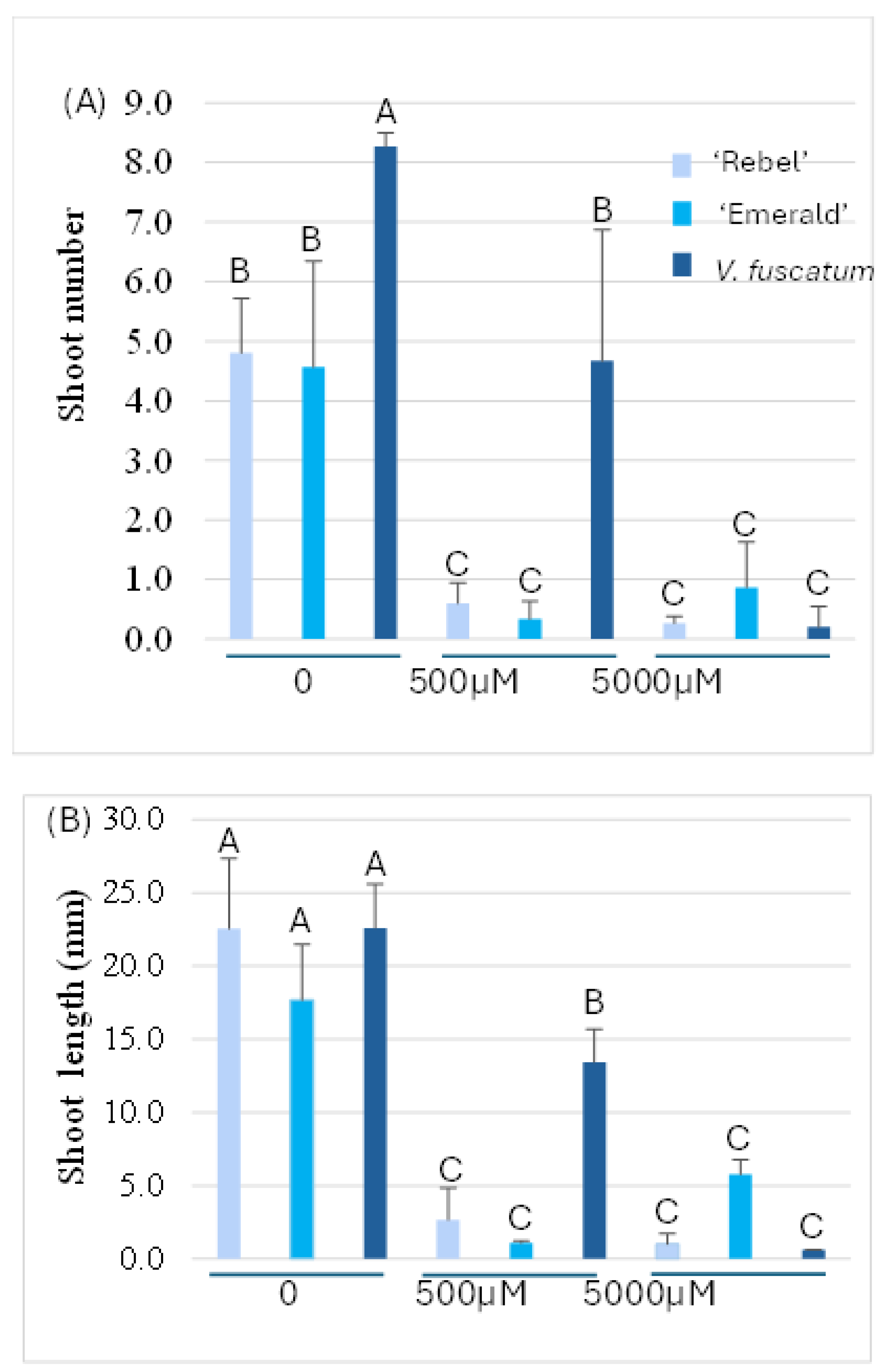
Figure 5.
Stem thickness of shoots micro-propagated from polyploidy-induced SHB (A) and V. fuscatum (B) compared to the respective mixoploid and wildtype controls. Measurement was performed after 100 days of transfer. Different letters on the top of the bar indicate statistical difference at p<0.05.
Figure 5.
Stem thickness of shoots micro-propagated from polyploidy-induced SHB (A) and V. fuscatum (B) compared to the respective mixoploid and wildtype controls. Measurement was performed after 100 days of transfer. Different letters on the top of the bar indicate statistical difference at p<0.05.
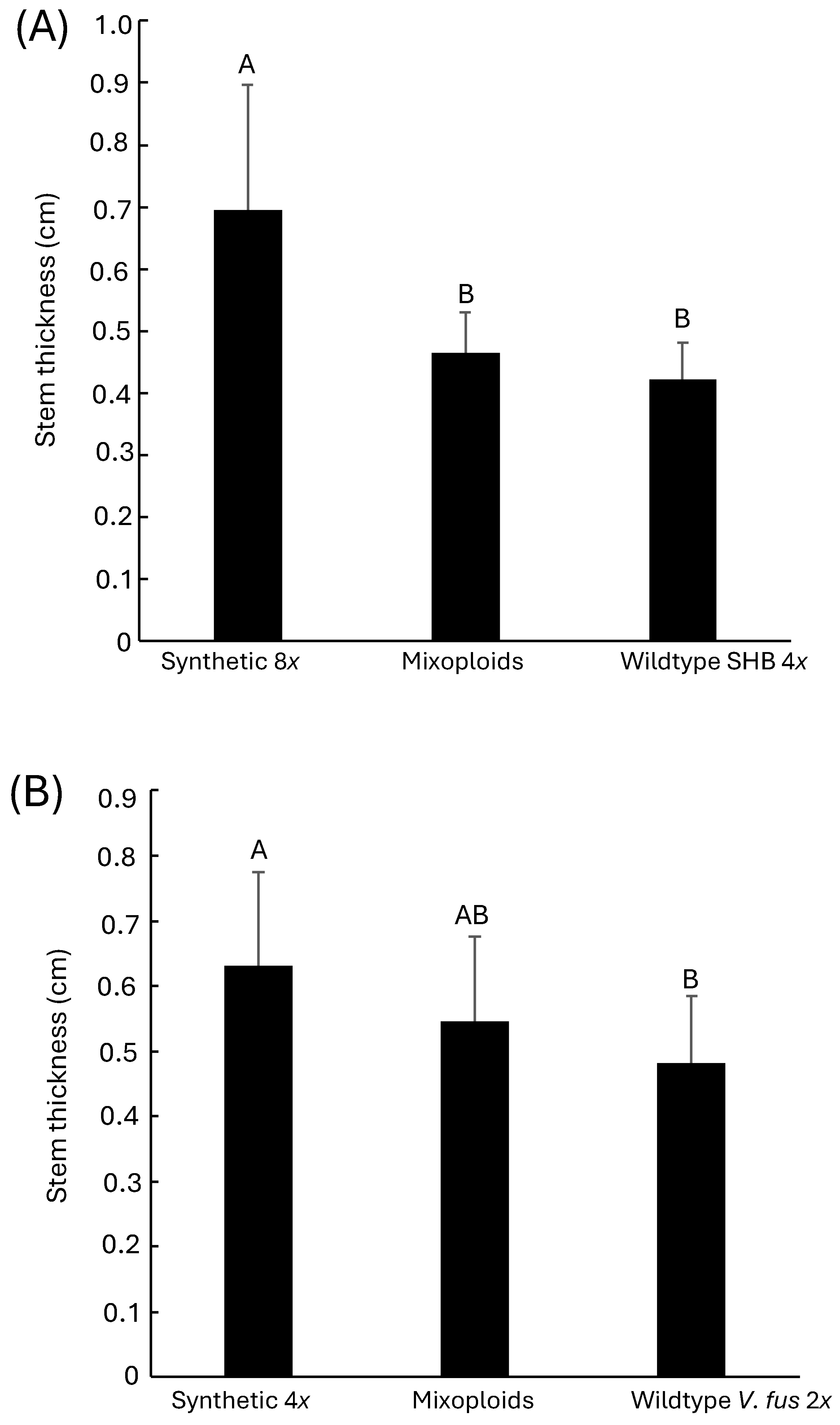
Figure 6.
Images of a diploid V. fuscatum wildtype plant (A), a synthetic tetraploid V. fuscatum plant (B), a tetraploid wildtype ‘Emerald’ plant (C), a synthetic octoploid ‘Emerald’ plant (D), a tetraploid wildtype ‘Rebel’ plant (E), and a synthetic octoploid ‘Rebel’ plant rooted from tissue cultured explants and grown in 1-gallon pots for eight months.
Figure 6.
Images of a diploid V. fuscatum wildtype plant (A), a synthetic tetraploid V. fuscatum plant (B), a tetraploid wildtype ‘Emerald’ plant (C), a synthetic octoploid ‘Emerald’ plant (D), a tetraploid wildtype ‘Rebel’ plant (E), and a synthetic octoploid ‘Rebel’ plant rooted from tissue cultured explants and grown in 1-gallon pots for eight months.

Figure 7.
Stomata impressions under microscope. (A) diploid V. fuscatum wildtype control; (B) synthetic tetraploid V. fuscatum; (C) tetraploid ‘Emerald’ wildtype control; (D) synthetic octoploid ‘Emerald’; (E) tetraploid ‘Rebel’ wildtype control; (F) synthetic octoploid ‘Rebel’. .
Figure 7.
Stomata impressions under microscope. (A) diploid V. fuscatum wildtype control; (B) synthetic tetraploid V. fuscatum; (C) tetraploid ‘Emerald’ wildtype control; (D) synthetic octoploid ‘Emerald’; (E) tetraploid ‘Rebel’ wildtype control; (F) synthetic octoploid ‘Rebel’. .
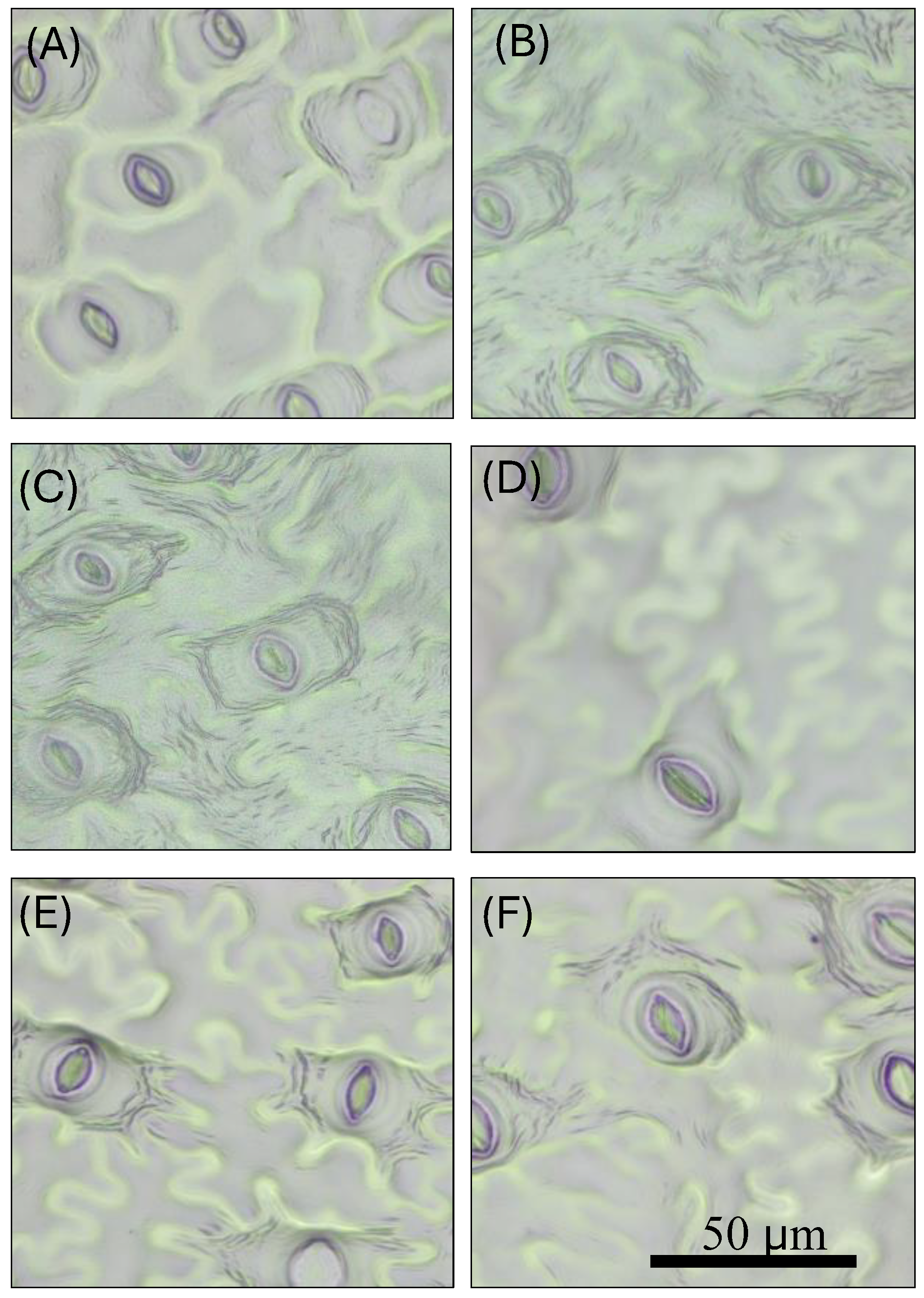

Table 1.
Ploidy levels of shoots regenerated from colchicine-treated nodal segments.
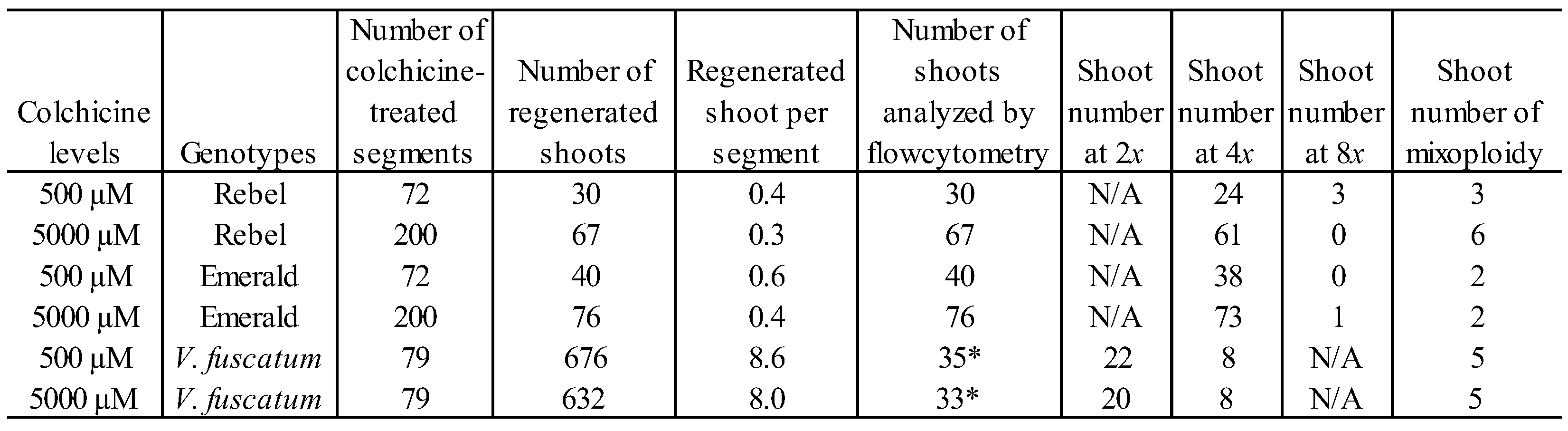
*Shoots with enlarged stem diameter were visually selected for flowcytometry analysis.
Table 2.
Plant morphological characteristics measured for wildtype and polyploidy-induced plants grown in 1-gallon pots. Different letters beside the numbers indicate statistical difference at p<0.05.
Table 2.
Plant morphological characteristics measured for wildtype and polyploidy-induced plants grown in 1-gallon pots. Different letters beside the numbers indicate statistical difference at p<0.05.
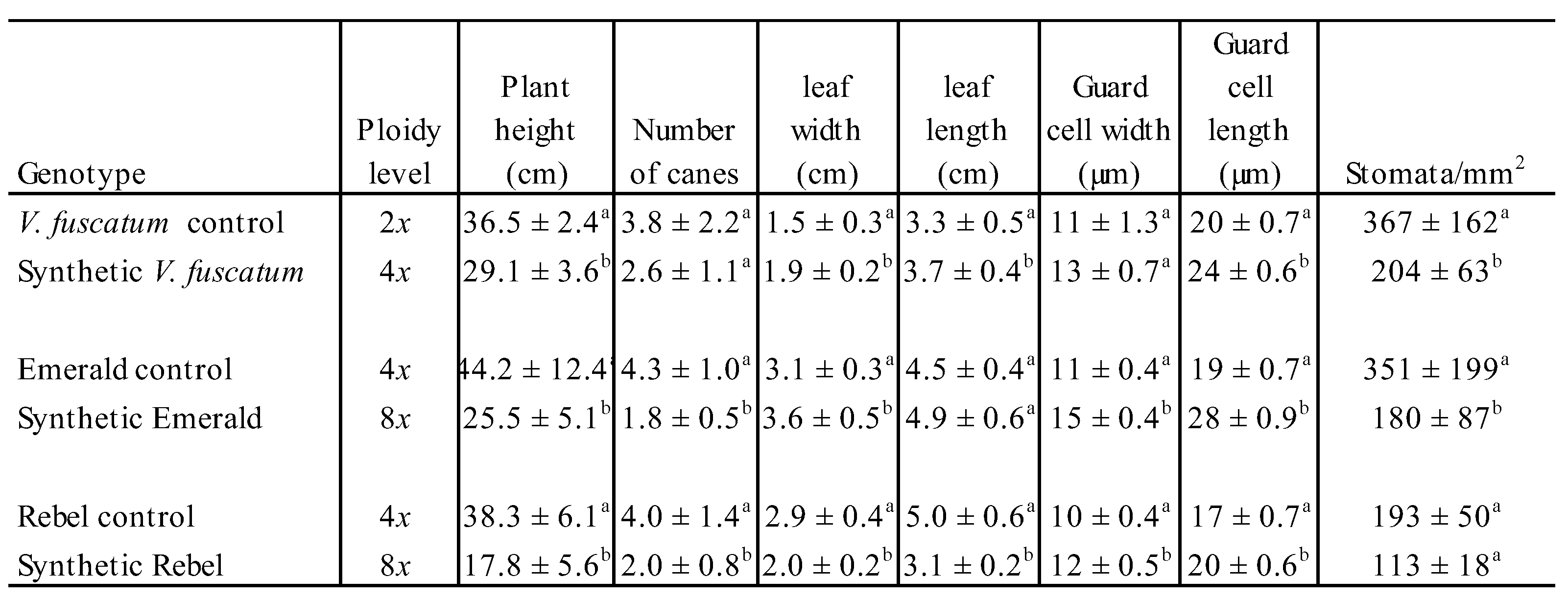
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



