
Submitted:
01 March 2025
Posted:
03 March 2025
You are already at the latest version
Abstract
Our paper conducted a re-examine of the Sauropod individuals PMoL from the Jehol Biota, Lower Cretaceous, Liaoning of China, including partial vertebrae and appendicula. First, we re-exam the osteology of the PMoL. Next, we used TNT 1.5 and PAUP 4.0 software and the newest matrix of the Titanosauriformes to analyze the phylogenetic position of the PMoL, the result revealed that PMoL was reconsidered in the Euhelopodid. Then, the comparison in the PMoL and other Titanosauriformes in the Jehol Biota was conducted. Finally, we conducted a paleobiogeographic analysis to infer the origination of the Euhelopodidae and considered that the origin region of the Euhelopodidae is Asia.
Keywords:
dinosaur
; Sauropod
; Euhelopodidae
; Titanosauriformes
; Titanosauria
1. Introduction
Titanosauriformes flourished in South America and Asia during the Cretaceous period [1]. In Asia, most taxa were excavated in China, until at the end of 2024, 32 taxa were recorded in China [2]. However, in the Jehol Biota, a famous biota for the Non-avian feathered Theropod dinosaur located in northeastern China of the Early Cretaceous [3], only three taxa of Titanosauriformes were recorded: Dongbeititan dongi, Liaoningotitan sinensis and Ruixinia zhangi [4,5,6].
The individual PMoL (PMoL: Paleontological Museum of Liaoning) is a Sauropod dinosaur individual that preserved partial cervical and dorsal vertebrae, scapula, ulna, and tibia. The individual was excavated in the Xiaobeigou Village, Beipiao City, Chaoyang City of Liaoning Province, China, close to the fossil point of the Liaoningotitan sinensis holotype [5]. However, this individual has been ignored for a long time, even some of its elements were wrongly classified as Liaoningotitan. To explore the Sauropod dinosaur in the Jehol Biota, we conducted total osteological research and phylogenetic analysis on the PMoL. We used The matrix in the TNT 1.5 and PAUP 4.0 software from Beeston et al., 2024, the newest matrix about the Titanosauriformes.
2. Materials and Methods
Systematic Paleontology
Saurischia [8]
Sauropodomorpha [9]
Sauropoda [10]
Titanosauriformes [11]
Somphospondyli [12]
Titanosauria [13]
Euhelopodidae [14]
Locality and horizon
The fossil point is in the Xiaobeigou Village, Shangyuan Town, Beipiao City, Chaoyang City, Liaoning Province, China (Figure 1). Yixian Formation, Aptian, Lower Cretaceous.
3. Results
3.1. Description
3.1.1. Cervical Vertebrae
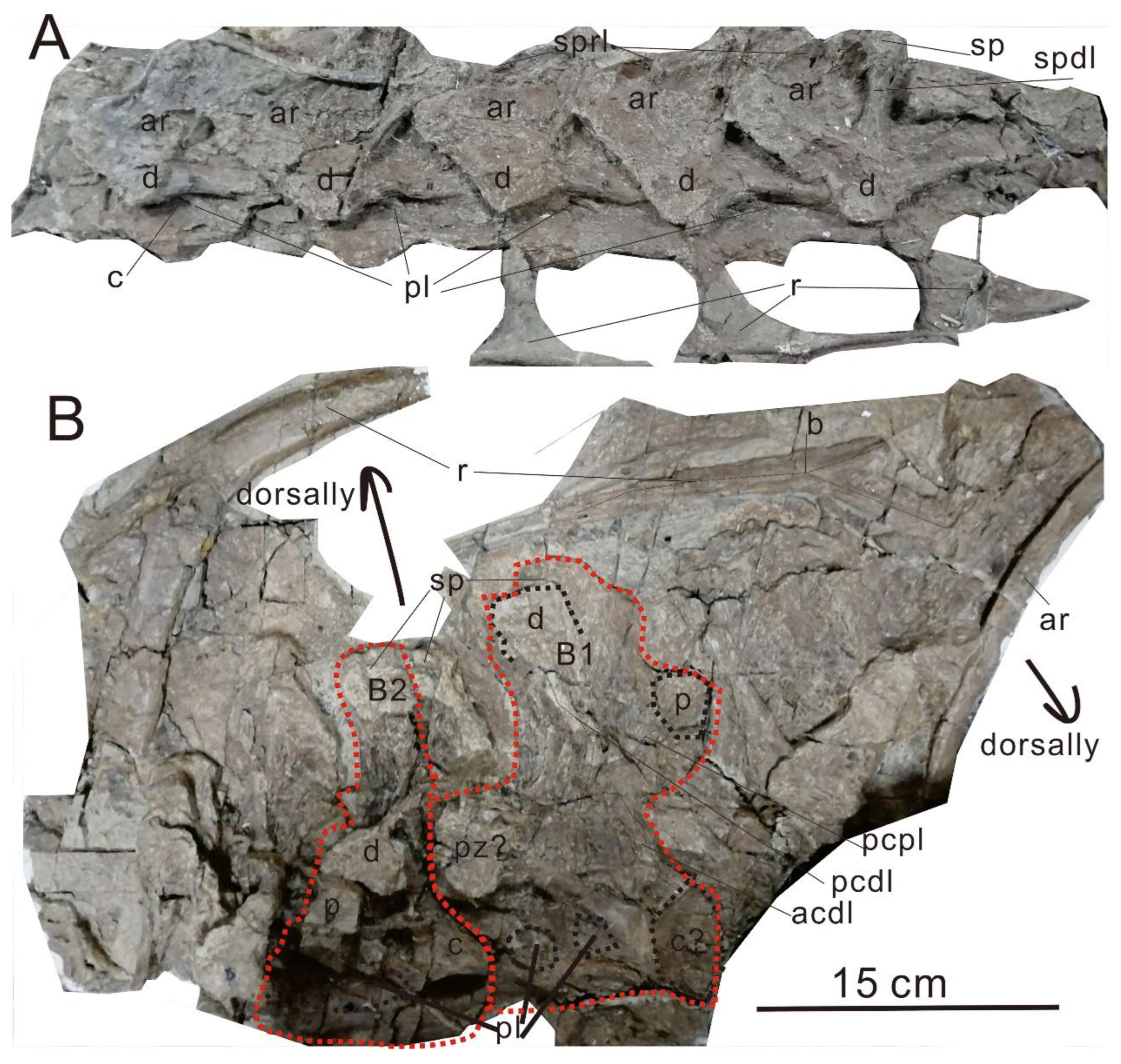
Five interrelated middle cervical vertebrae and one hindmost cervical vertebra are preserved. All middle cervical vertebrae are flat, presumably flattened by the rock bed. All neural arches and neural spines are incomplete. The height of the neural spines is increased progressively. The diapophyses are triangular and extend laterally. The diapophysis is elongated more than half the length of the centrum. Only the last three cervical ribs are well preserved and are double-head type. The cervical ribs are elongated. A lateral pneumatic fossa/foramen extends almost to the posterior end of the centrum, leaving only a thin strip of bone Posteriormost cervical and anteriormost dorsal neural spines. The spinoprezygaphyseal lamina is broken. The location of the cervical rib is on a level that is similar to that in the centrum. The lateral surface of the cervical rib developed a bulge. The location of the cervical ribs is aligned to the centrum. The angle between spinodiapophyseal lamina and the arch is acute. The dorsoventral height divided by the posterior centrum height of the hindmost cervical vertebra is greater than 1.0. The hindmost cervical vertebra developed a long neural arch.
3.1.2. Dorsal Vertebrae
Four foremost dorsal vertebrae are preserved, but only two vertebrae were preserved well (B1 and B2), and the orientation of the dorsal vertebrae is inverted with the hindmost cervical vertebra. The neural spine is not bifurcated and vertical to the centrum. The centrum is opisthocoelous. In B1, the neural spine is vertical to the vertebra. The diapophysis extends dorsoposteriorly. The parapophysis is extending dorsoanteriorly. The anterior centrodiapophyseal lamina, posterior centrodiapophyseal lamina, and posterior centroparapophyseal lamina consist of a K form. The pleurocoel is divided by a short plate.
In B2, the diapophysis and parapophysis are vertical to the centrum. A shallow and long pneumatic (pleurocoel) foramen is on the dorsal vertebra's lateral side. The type of the dorsal rib is plate-like.
Figure 2.
Vertebrae of PMoL. Scale bar: 15 cm. A: middle cervical vertebrae in left view; B: posterior vertebra and anterior dorsal vertebrae in right view. Abbreviations: acdl, anterior centrodiapophyseal lamina; ar, arch; b, bulge; c, centrum; d, diapophysis; p, parapophysis; pcdl, posterior centrodiapophyseal lamina; pcpl, posterior centroparapophyseal lamina; pl, pleurocoel; r, rib; sp, spine; spdl, spinodiapophyseal lamina; sprl, spinoprezygaphyseal lamina.
Figure 2.
Vertebrae of PMoL. Scale bar: 15 cm. A: middle cervical vertebrae in left view; B: posterior vertebra and anterior dorsal vertebrae in right view. Abbreviations: acdl, anterior centrodiapophyseal lamina; ar, arch; b, bulge; c, centrum; d, diapophysis; p, parapophysis; pcdl, posterior centrodiapophyseal lamina; pcpl, posterior centroparapophyseal lamina; pl, pleurocoel; r, rib; sp, spine; spdl, spinodiapophyseal lamina; sprl, spinoprezygaphyseal lamina.
3.1.3. Scapula
Only one right scapula and partial coracoid are preserved and the medially of all individuals are covered by gypsum, therefore, only the laterally of the scapula and the coracoid are visible. The scapula is 100 cm in length. The dorsal side is thick, and the ventral side is thin. The proximal end of the scapula is medially curved, similar to that in Somphospondyli. The ratio of the maximum dorsoventral height to the minimum dorsoventral height of the scapular blade is 1.7, less than that in the Daxiatitan binglingi and Jiangshanosaurus lixianensis but greater than that in the Yongjinglong datangi and Patagotitan mayorum [15,16,17,18,[15,16,17,18,]. The orientation of the scapula blade long-axis concerning coracoid articulation is 72°. The scapular glenoid face is medially, dissimilar from the Jiangshanosaurus and Liaoningotitan [16]. An anteroposteriorly concave region posterior to the scapular glenoid, followed by a flattened area. The scapula has a lateral ridge in the middle of the shaft and extends to the anterior and posterior, similar to that of Jiangshanosaurus lixianensis and Patagotitan mayorum [16,18], and speculated to be the attachment point of the subcoracoscapularis muscle. The ratio of the overall length of the scapula to its narrowest dorsoventral length is 6.13, less than that in the Daxiatitan binglingi [15]. The acromion process is not preserved. The posterior margin of the acromial plate is flat. The acromial ridge is developed but broken. The angle between the acromion posterior region and the scapular shaft long axis is 32°. The dorsal margin of the acromion is broken, therefore we cannot estimate whether there is an acromion corner or not. The scapula developed no subtriangular process in the anteroventral corner of the scapular blade, which is the tuberosity (located in the anterior ventral side of the scapula blade, also called the attachment point of the triceps brachii muscle), dissimilar to that in Yongjinglong datangi and Daxiatitan binglingi [17,15]. The scapular suture is vertical to the coracoid almost. The thickest area of the shaft is located in the 1/3 area of the blade. The dorsoventral height of the distal end is greater than the dorsoventral height of the proximal end of the blade. The distal end extends to the dorsoposterior and posterior side, with an attachment point of the teres major muscle (tm) located on the laterally distal end of the scapula.
3.1.4. Ulna
The right ulna is preserved, triradiate (Figure 3), 45 cm long, and only the posterior side is visible. The width of the proximal end is 18 cm, and the width of the distal end is 14 cm.
3.1.5. Tibia
The proximal end extends lateromedially. The cnemial crest is well-developed and extends anterolaterally to the 1/3 position of the shaft. The crest length is 30% of the width of the tibial proximal end. The lateral margin of the cnemial crest in the anterior view is subtriangular, forming a point. The fibular articular surface is flat. The length of the proximal end/middle position is 2.04. The narrowest position of the tibia is located at the distal end. The distal end extends to both anteriorly and posteriorly and the length is 50% of the length of the proximal end. The second cnemial crest is absent, similar to that in Euhelopodidae [7]. The proximal lateral face with a double ridge extending distally from the lateral projection of the proximal articular area and posterolateral fossa posterior to the double ridge, containing a lower tuberosity and an upper deep pit is absent. The location of the anterior process is higher than that of the posterior process. The appendicula is shown in Figure 3.
3.2. Phylogenetic Analysis
Our analysis used the matrix modified from [19] in TNT 1.5 software and PAUP 4.0 [20] for the phylogenetic analysis, in which the matrix includes 131 taxa (OTUs) and 560 characteristics. We added PMoL and Liaoningotitan into the matrix. Extended implied weighting (EIW) analyses were used with the following settings: max.trees was set to 10, 000; tree bisection and reconnection (TBR) was used; new technology search was selected; random addition sequences were changed from 1 to 1,000 addseqs; sect search, ratchet, drift, and tree fusing were all used; set K=9, similar in the previous analysis [16,19]. All other options were set to default. The results show 3 strict consensus trees, a tree length of 2,840, a CI of 0.208, and a RI of 0.584. Standard bootstrap with the number of replicates changed from 100 to 5,000. The final result identifies that Liaoningotitan is a member of the Euhelopodidae, and PMoL is a member of the Euhelopodidae too, as shown in Figure 4.
4. Discussion
4.1. The phylogenetic Position of the Titanosauriformes in China
In the result of our analysis, Titanosauriformes in China reflected a high degree of diversity. Fusuisaurus was reconsidered in the Basal Titanosauriform, similar to that of the initial analysis [21]. Liubangosaurus was reconsidered as a Non-Neosauropod taxon and a Lithostrotian in the previous analyses and as a Euhelopodid in our analysis [23,7]. Qiaowanlong was classified as a Brachiosaurid in the initial analysis too [22]. The Dongbeititan is considered a Basal-Somphospondylan. Liaoningotitan, Yongjinglong, Jiangshanosaurus, Qiaowanlong, Liubangosaurus, Euhelopus, Gobititan, two Huanghetitan taxa, and PMoL are reconsidered Euhelopodidae. Daxiatitan, Ruyangosaurus, Xianshanosaurus, Huabeisaurus, Dongyangosaurus and Mongolosaurus are Titanosaurian. In which, Dongyangosaurus and Mongolosaurus are reconsidered as Lithostrotian.
4.2. Euhelopodid in the Jehol Biota
At the end of 2024, three Sauropod taxa were named in the Jehol Biota: Dongbeititan dongi, Liaoningotitan sinensis, and Ruixinia zhangi, all taxa were considered as the Euhelopodid exclude Dongbeititan (this paper; [6]). In addition, some fragmentary individuals reveal the Euhelopodid existence in the Jehol Biota, such as teeth [24], indicating that the Jehol Biota is an important habitant of Euhelopodid dinosaurs. PMoL increased the diversity of the Euhelopodid in the Jehol Biota. PMoL is different from other Sauropods from the Jehol Biota in the following positions: 1. The location of the cervical ribs is aligned to the centrum, dissimilar to that of the Liaoningotitan and Ruixinia. 2. The ratio of the length of the proximal end to the middle position of the PMoL is 2.04, greater than that of the Dongbeititan (1.89) and less than that of the Liaoningotitan (2.7) and Ruixinia (2.58) [4, 6], support that the PMoL is an independent taxon that different to others.
4.3. The Paleobiogeographic Reconstruction of Euhelopodidae
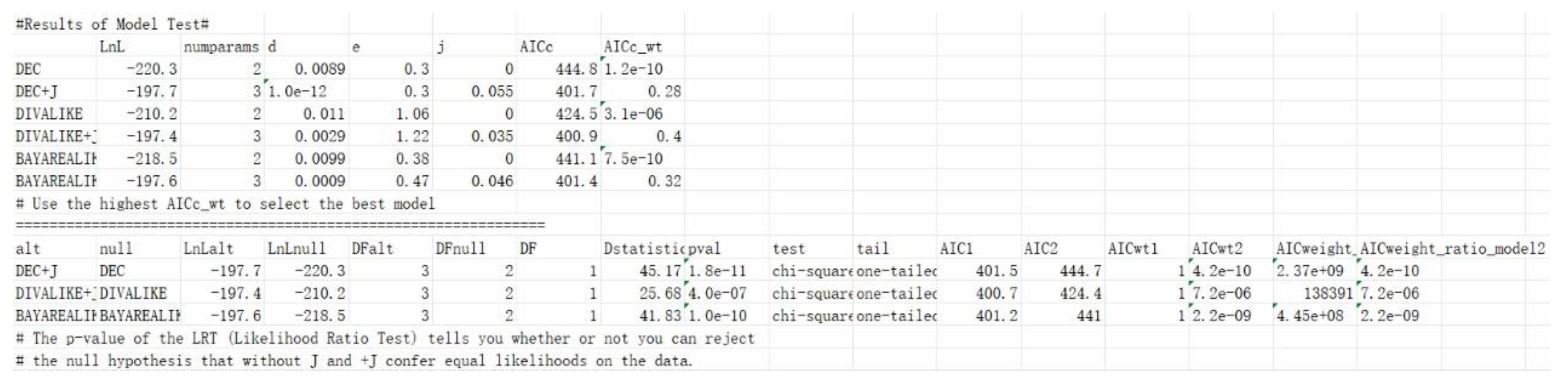
This study utilized the Rasp software R package BioGeoBears [25] to reconstruct the ancestral distribution of Titanosauriformes in Asia. The BioGeoBears was used to add a time axis to the tree, root length =2 Ma (Million years). FAD (First Appearance Date) and LAD (Last Appearance Date/Extinct Date) of taxa were referred to the Paleobiology Database (https://www.paleobiodb.org/) and previous research [1], the geological period information refers to the 2020 version of the International Commission on Stratigraphy Chronostratigraphic Chart. BioGeoBears supports DEC, DEC + J, DIVA, DIVA + J, DIVALIKE, DIVALIKE+J, BAYAREALIKE, and BAYAREALIKE + J models to analyze the ancestral distribution of creatures. These models allow different vicariance to occur on nodes. In the analysis, we put the potential ancestral distribution into geographical areas A – C and allow each ancestor to occupy up each two geographical areas (such as AB, AC, etc). By comparing all models, we chose model BAYAREALIKE for our analysis, the result of reconstruction is seen in Figure 5, and the model test result is seen in Figure 6. The disk represents the proportion of the possibility of originating from somewhere.
Our result revealed The Euhelopodidae originated from Asia but is not an endemic taxon in Asia. The Euhelopodidae consists of a gigantic monophyletic group. The group has a spread distribution range including Asia, Europe, and South America, but excluding North America, Africa, and Australia. In our reconstruction, the Euhelopodid origin is estimated in the Berriasian, which is compatible with the earliest Euhelopodid known-Euhelopus zdanskyi. The MRCA (the most common ancestor) of European and South American taxa possibly emigrated out from Asia by land bridge in the Early Cretaceous, the origination of them is estimated to generally correspond to the Asia and Europe separated by a massive transgression in the Early Cretaceous.
Most taxa remain in Asia. Asian Euhelopodid is divided into two Monophyletic groups and reveals an obvious regional distribution difference between North and South. All taxa in the upper blade are found in northern Asia excluding Jiangshanosaurus. All taxa in the lower blade are found in southern Asia excluding Qiaowanlong, indicating that a massive spread event appeared in the Asia of the Early Cretaceous, it is also speculated that it is also related to the geographical isolation caused by transgression or other geological phenomenon, we prospect the geological and paleontological evidence excavated in the future.
5. Conclusions
Our paper researched and analyzed an incomplete Sauropod individual PMoL from the Yixian Formation of the Jehol Biota and considered the PMoL as a possible Euhelopodid, and considered that the origin region of the Euhelopodidae in Asia by the paleobiogeographic reconstruction. However, we also considered that the Euhelopodidae is not an endemic taxon in Asia, they may also be distributed in Europe and South America. This viewpoint may have rewritten the evolutionary history of the Euhelopodidae. However, we should also note that the poor preservation of the individual and fossil record has limited our ability to conduct further research on it. Therefore, we look forward to seeing more complete Sauropod dinosaur fossils excavated in the future to solve these problems.
Supplementary Materials
The supplementary materials ‘FileS2.tnt.txt’ can be found and used at https://figshare.com/account/items/28517621/edit.
Author Contributions
All works were conducted by only one author Mr Shan bingqing. Paleontological Museum of Liaoning, Shenyang Normal University supplied fossil individual.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Acknowledgments
I am grateful to all teachers and staff from the Paleontological Museum of Liaoning, Shenyang Normal University for assistance and best wishes to them.
Conflicts of Interest
The author declares no conflict of interest.
References
- Gorscak, E. ; O‘Connor, P, M. Biology Letters 2016, 1–5. [Google Scholar] [CrossRef]
- Han, F.; Yang, L.; Lou, F.; Corwin, S.; Xu, X.; Qiu, W.; Liu, H.; Yu, J.; Wu, R.; Ke, Y.; Xu, M.; Hu, J.; Lu, P. A new titanosaurian sauropod, Gandititan cavocaudatus gen. et sp. nov., from the Late Cretaceous of southern China. Journal of Systematic Palaeontology 2024, 22:1-23. [CrossRef]
- Zhang, L. Analysis on fossil community of tetrapods in Lujiatun bed of Lower Cretaceous Yixian Formation in Beipiao, Liaoning. Global Geology 2020, 39, 738–744. [Google Scholar]
- Wang, X.; You, H.; Gao, C.; Cheng, X.; Liu, J. Dongbeititan dongi the first Sauropod Dinosaur from the Lower Cretaceous Jehol Group of Western Liaoning Province China. Acta Geologica Sinica 2007, 6, 911–916. [Google Scholar]
- Zhou, C.; Wu, W.; Sekiya, T.; Dong, Z. A New Titanosauriformes dinosaur from Jehol Biota of western Liaoning China. Global Geology 2018, 37, 328–333. [Google Scholar] [CrossRef]
- Mo, J.; Ma, F.; Yu, Y.; Xu, X. A New Titanosauriform Sauropod with An Unusual Tail from the Lower Cretaceous of Northeastern China. Cretaceous Research 2022: 1-51. [CrossRef]
- Mannion, P, D. ; Upchurch, P.; Barnes, R, N.; Mateus, O. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriformes. Zoological Journal of the Linnean Society 2013, 168, 98–206. [Google Scholar]
- Seeley, H, G. On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London 1887, 43, 165–171. [Google Scholar]
- Huene, V, F, F. Die fossil Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte. Monographien zur Geologie und Palaeontologie 1932, 1, 1–361.
- Marsh, O, C. Principal characters of American Jurassic dinosaurs. Pt. I. American Journal of Science 1878, 16:411-416. [CrossRef]
- Salgado, L.; Coria, R, A. ; Calvo, J, O. Evolution of titanosaurid sauropods. 1: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 1997, 34, 3–32. [Google Scholar]
- Wilson, J, A.; Sereno, P. Early Evolution and Higher-Level Phylogeny of Sauropod Dinosaurs. 1998. [CrossRef]
- Bonaparte, J, F. ; Coria, R, A. Un nuevo y gigantesco saurópodo titanosaurio de la Formación Río Limay (Albiano–Cenomaniano) de la Provincia del Neuquén, Argentina. Ameghiniana 1993, 30, 271–282. [Google Scholar]
- A, S, Romer. Osteology of the Reptiles, University of Chicago Press 1956: 1-772.
- You, H.; Li, D.; Zhou, L.; Ji, Q. Daxiatitan binglingi: a Giant Sauropod Dinosaur from The Early Cretaceous of China. Gansu Geology 2008, 17, 2–10. [Google Scholar]
- Mannion, P, D.; Upchurch, P.; Jin, X.; Zheng, W. New information on the Cretaceous sauropod dinosaurs of Zhejiang Province China: impact on Laurasian Titanosauriformes phylogeny and biogeography. Royal Society Open Science 2019, 1-22. [CrossRef]
- Li, L.; Li, D.; You, H.; Dodsen, P. A New Titanosauria Sauropod from the Hekou Group (Lower Cretaceous) of the Lanzhou-Minhe Basin, Gansu Province, China. PLoS ONE 2014, 9, e85979. [Google Scholar] [CrossRef]
- Otero, A.; Carballido, J, L.; Moreno, P, A. The appendicular osteology of Patagotitan mayorum (Dinosauria, Sauropoda). Journal of Vertebrate Paleontology 2020, 1-20. [CrossRef]
- Beeston, S, L.; Poropat, S, F.; Mannion, P, D.; Pentland, A, H.; Enchlmaier, M, J.; Sloan, T.; Elliott, D, A. Reappraisal of sauropod dinosaur diversity in the Upper Cretaceous Winton Formation of Queensland, Australia, through 3D digitization and description of new specimens. PeerJ 2024 1-105. [CrossRef]
- Goloboff, P.; Catalano, S. TNT, version 1.5, with a full implementation of phylogenetic morphometrics. Cladistics 2016. [CrossRef]
- Mo, J.; Wang, W.; Huang, Z.; Huang, X.; Xu, X. A Basal Titanosauriform from the Early Cretaceous of Guangxi, China. Acta Geologica Sinica 2006, 80, 486–489. [Google Scholar]
- Li, D.; You, H. The first well-preserved Early Cretaceous Brachiosaurid dinosaur in Asia. Proceedings of the Royal Society B Biological Science 2009, 276, 4077–4082. [Google Scholar]
- Mo, J.; Xu, X.; Buffetaut, E. A New Eusauropod dinosaur from the Lower Cretaceous of Guangxi Province, Southern China. Acta Geologica Sinica (English edition) 2010, 84, 1328–1335. [Google Scholar]
- Zhang, H.; Yin, Y.; Pei, R.; Zhou, C. Early-diverging Titanosauriform (Dinosauria, Sauropoda) Teeth from the Lower Cretaceous Yixian Formation of Southeastern Inner Mongolia, Northeast China. Acta Geologica Sinica (English Edition) 2024, 98: 303– 310. [CrossRef]
- Yu, Y.; Harris, A, J.; Blair, C.; He, X, J. RASP (Reconstruct Ancestral State in Phylogenies): a tool for historical biogeography. Molecular Phylogenetics and Evolution 2015, 87, 46–49. [Google Scholar] [PubMed]
Figure 1.
Geographic of PMoL. Locality of PMoL (red point) in Xiaobeigou Village, Shangyuan Town, Beipiao City, Chaoyang City, Liaoning Province, Map copyright: https://map.tianditu.gov.cn/.
Figure 1.
Geographic of PMoL. Locality of PMoL (red point) in Xiaobeigou Village, Shangyuan Town, Beipiao City, Chaoyang City, Liaoning Province, Map copyright: https://map.tianditu.gov.cn/.
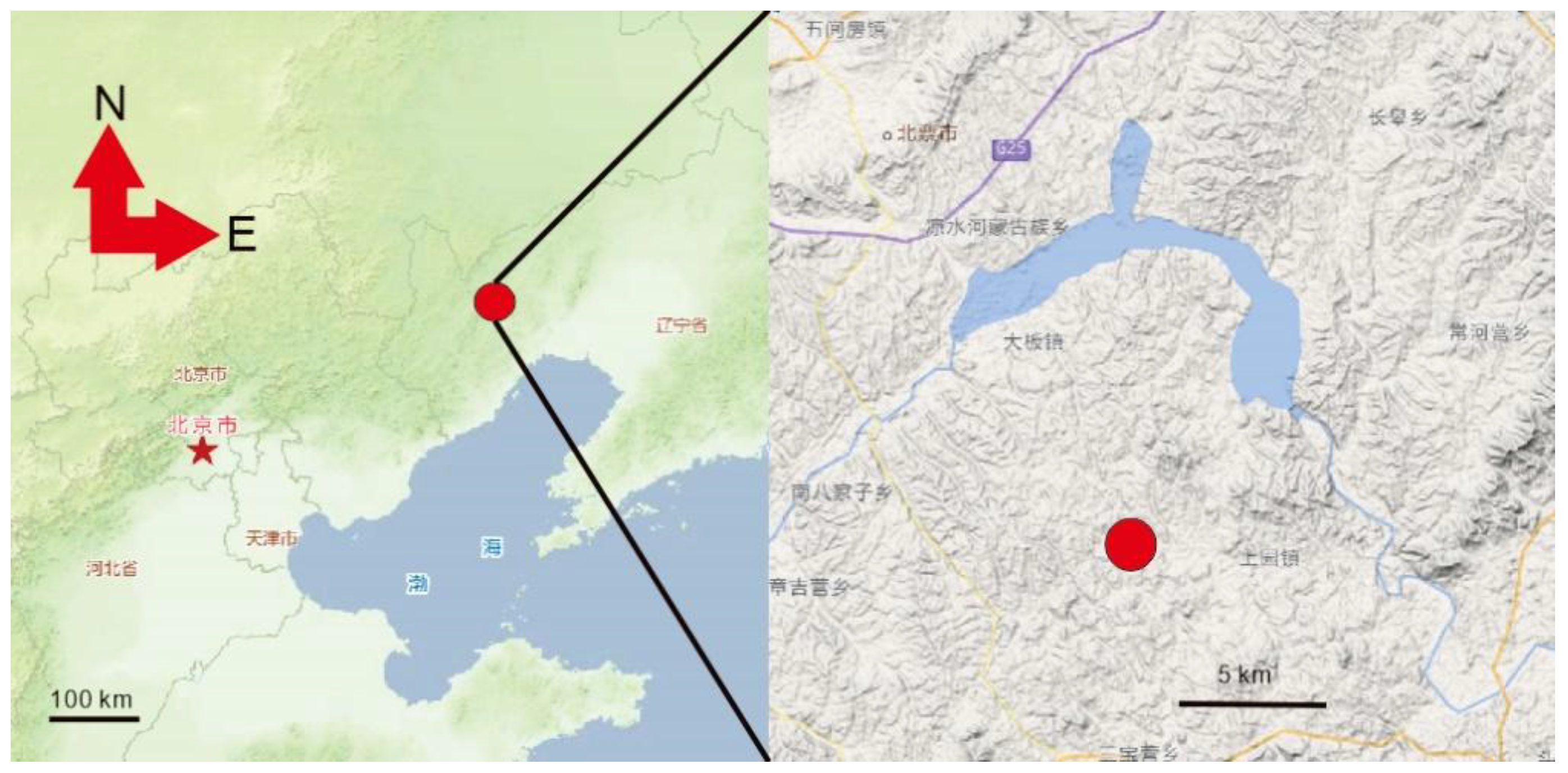
Figure 3.
Appendicular elements of PMoL. Scale bar: 15 cm. A: Scapula in lateral view. B: Tibia in anterior view. C: Ulna in medial view. Abbreviations: acr acromial ridge; ap: anterior process; cc: cnemial crest; gl, glenoid lip; ltr, lateral ridge; pp: posterior process; scb, scapular blade; tm, the attachment point of the teres major muscle.
Figure 3.
Appendicular elements of PMoL. Scale bar: 15 cm. A: Scapula in lateral view. B: Tibia in anterior view. C: Ulna in medial view. Abbreviations: acr acromial ridge; ap: anterior process; cc: cnemial crest; gl, glenoid lip; ltr, lateral ridge; pp: posterior process; scb, scapular blade; tm, the attachment point of the teres major muscle.
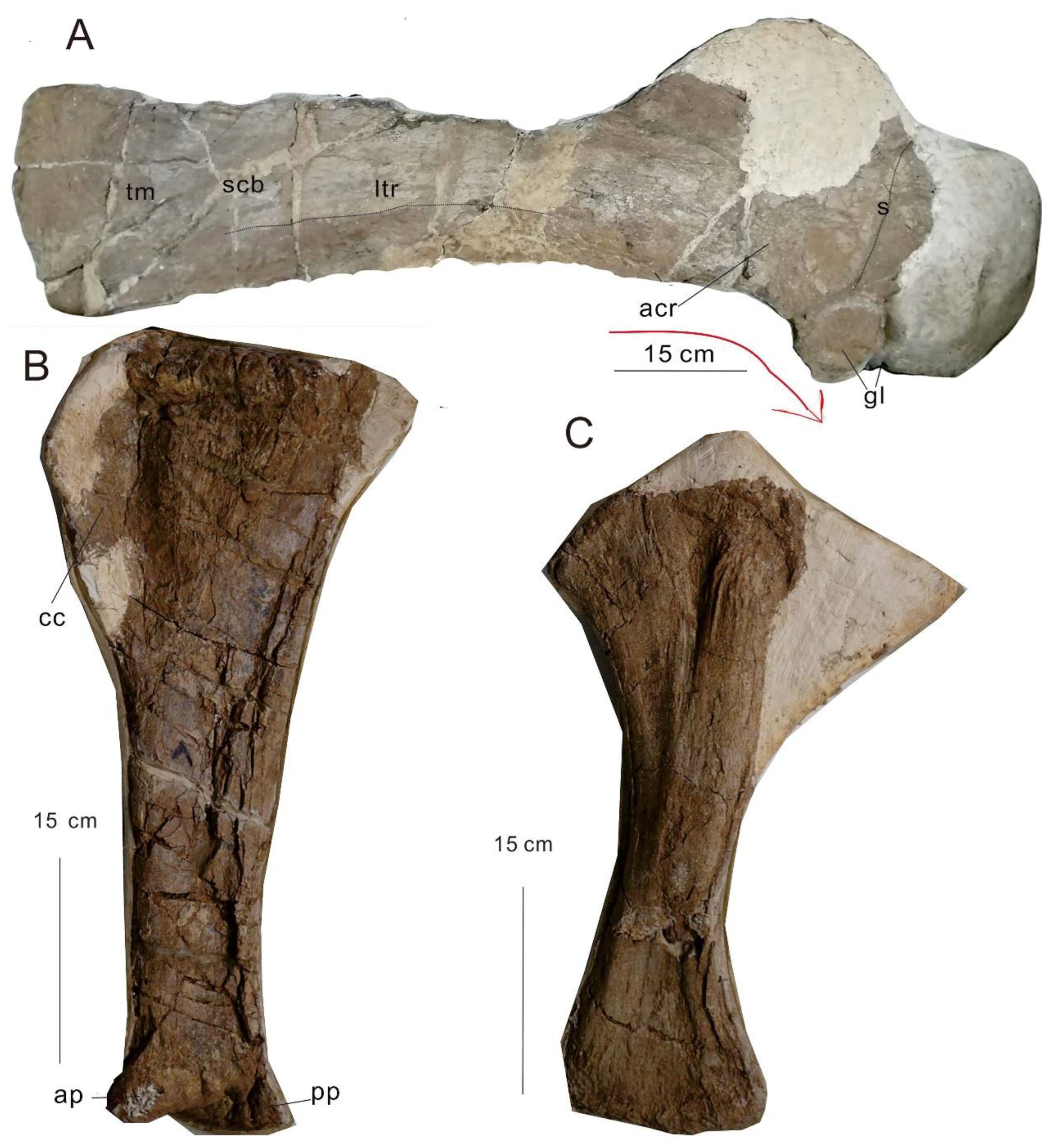
Figure 4.
Phylogenetic analysis of strict consensus tree of PMoL (red arrow). Matrix from [19].
Figure 4.
Phylogenetic analysis of strict consensus tree of PMoL (red arrow). Matrix from [19].
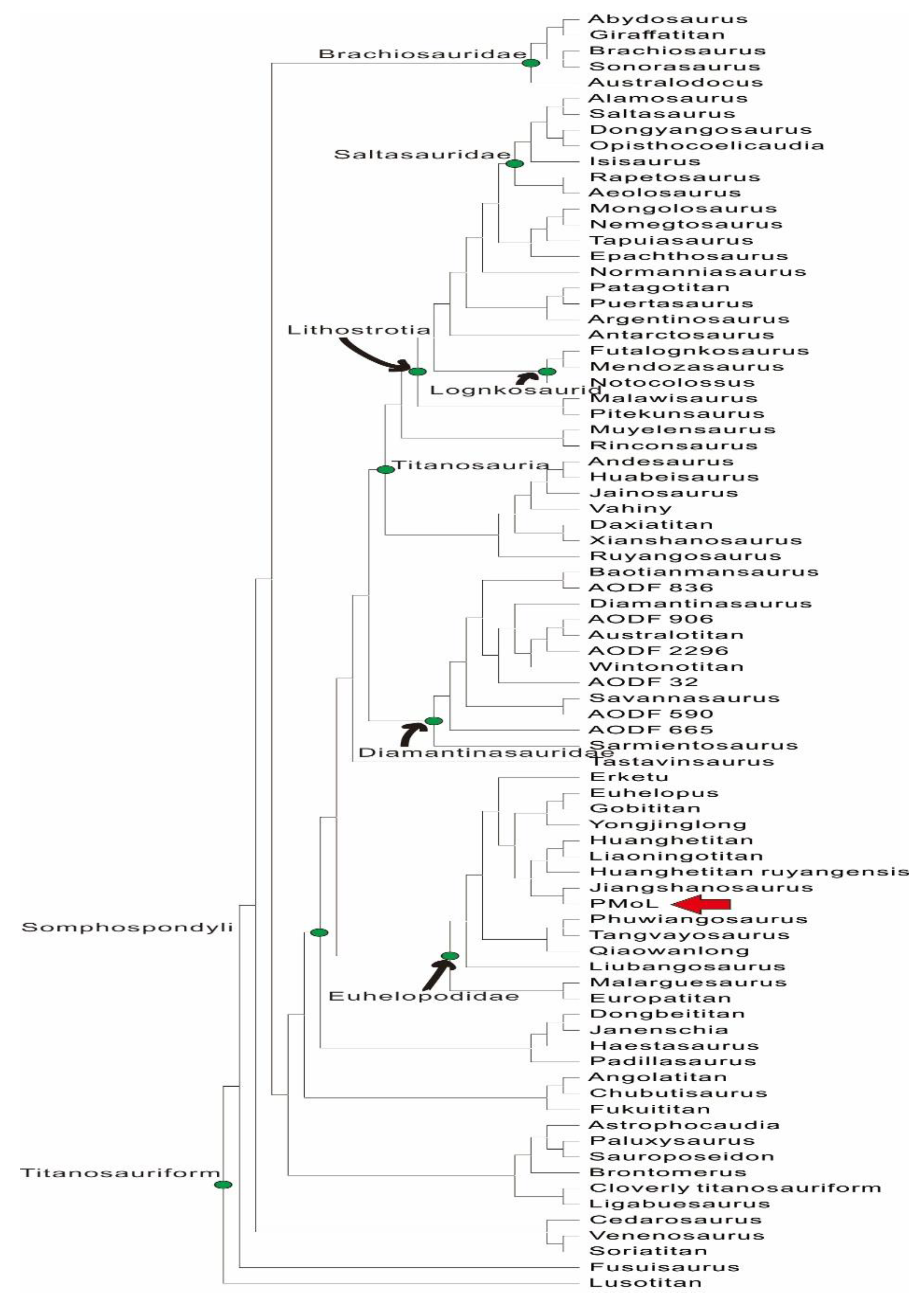
Figure 5.
The paleobiogeographic reconstruction of Euhelopodidae (including PMoL and Liaoningotitan).
Figure 5.
The paleobiogeographic reconstruction of Euhelopodidae (including PMoL and Liaoningotitan).
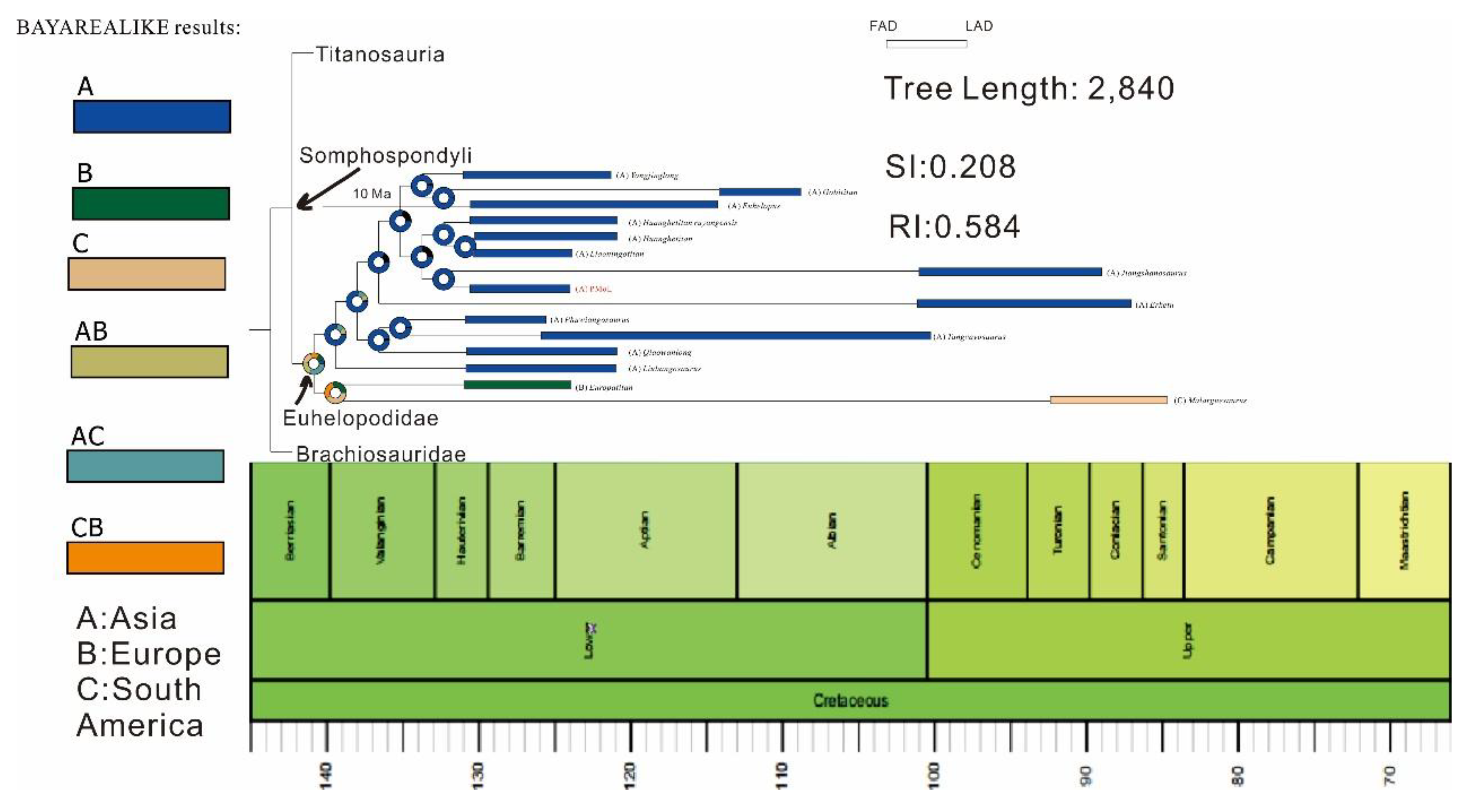
Figure 6.
Model test results of the paleobiogeographic reconstruction of Euhelopodidae (including PMoL and Liaoningotitan).
Figure 6.
Model test results of the paleobiogeographic reconstruction of Euhelopodidae (including PMoL and Liaoningotitan).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



