
Submitted:
13 January 2025
Posted:
14 January 2025
You are already at the latest version
Abstract
Black Soldier Fly Larvae (BSFL), Hermetia illucens, present a transformative approach to sustainable aquaculture and waste management by converting organic waste into nutrient-rich biomass. This study highlights BSFL-derived products as a sustainable alternative for aquafeeds, delivering essential nutrients and significant concentrations of bioactive and antioxidative polyphenols. We evaluated five organic waste types—Kitchen Waste (KW), Agricultural Waste (AW), Aquaculture Sludge (AS), Aquaculture Offal (AO), and a Mixed diet (MX)—assessing their impact on BSFL growth, Feed Conversion Ratio (FCR), bioconversion, Protein Efficiency Ratio (PER), survival, and nutritional composition, including Total Polyphenol Concentration (TPC). This is the first report to document and quantify the transfer of polyphenols from waste substrates to larvae, showing that larvae fed plant-based feeds contained significantly higher (5.8 - 7.4 mg GAE/100g) TPC than those fed with an animal-based feed (0.60 - 0.9 mg GAE/100g) (p < 0.05). Our findings emphasize the potential of BSFL to integrate waste valorization into circular economy frameworks, reduce organic waste, and enhance aquafeeds’ functional properties, contributing to ocean sustainability and the global transition to more sustainable aquaculture practices.
Keywords:
Sustainable Aquaculture
; Circular Economy
; Waste Valorization
; Organic Waste Recycling
; Functional Feed Additive
; Bioactive Polyphenols
; Black Soldier Fly Larvae
1. Introduction
As global aquaculture continues to expand to meet the growing demand for fish and seafood, there is increasing pressure to develop sustainable feed sources or for the industry to become more sustainable. Fishmeal and soybean meal have been and are the primary protein ingredients in aquaculture diets. However, concerns over the environmental impact of producing fishmeal, its fluctuating prices, and limited availability, has prompted the search for alternative feed ingredients [1,2]. On the other hand, inclusion of soybean-based proteins at an excessive level have been found to cause negative effects on salmon intestinal morphology and gut microbiota [3]. An alternative to fish and soybean meal is black soldier fly larvae meal (BSFLM), which has gained significant attention not only as a protein source but also as a potential functional feed additive due to its ability to accumulate and produce beneficial nutrients when reared on specific organic waste [4,5].
Black soldier fly larvae (Hermetia illucens) (BSFL) are remarkable bio-converters, capable of transforming a wide variety of organic waste into nutrient-dense biomass. These larvae efficiently upcycle nutrients from diverse waste streams, including agro-industrial by-products, food waste, and manure, converting them into valuable and nutrient-rich products. The larvae can not only accumulate proteins and lipids, but also other bioactive compounds such as polyphenols—plant-based secondary metabolites known for their antioxidant properties [6,7]. Polyphenols have been shown to reduce oxidative stress in fish, potentially enhancing immune function and overall performance [8]. The ability of BSFL to accumulate polyphenols and other bioactive compounds highlights the potential of BSFLM not just as a protein ingredient but as a functional feed additive that could improve fish health through aquaculture diets [5].
Despite these promising qualities, a significant barrier to the widespread adoption of BSFLM as a primary protein source is its current market price, which remains considerably higher than traditional protein sources like fishmeal and soybean meal [9]. However, due to the unique nutritional and functional properties of BSFLM, it holds great potential as a feed additive rather than a complete or partial replacement for conventional protein sources [10].
Moreover, the process of upcycling organic waste into high-value biomass through BSFL aligns with the principles of a circular economy, where waste is diverted from landfills and repurposed into valuable products. This bioconversion not only reduces the environmental burden of waste disposal but also creates a nutrient-rich product that can support more sustainable aquaculture practices [11]. BSFL can process a variety of low-value organic wastes, transforming them into high-quality biomass rich in proteins, fats, and bioactive compounds, thereby contributing to the sustainability of both waste management and aquaculture sectors [4]. Research has increasingly demonstrated the additional benefits of BSFLM beyond its nutrient profile, particularly in enhancing gut health and microbial diversity in fish. Studies have shown that BSFLM can positively influence gut morphology microbiota, improve nutrient digestibility, and boost immune responses in species such as sea bream, meagre, and salmon [12,13,14]. These effects are attributed not only to the protein and fat content of BSFLM but also to the presence of bioactive compounds, which may play a role in modulating the gut microbiota and improving overall fish health [8]. These attributes reinforce the potential of BSFLM as a functional feed additive in aquaculture, enhancing its appeal as a sustainable option for improving fish health and production efficiency.
In this study, we investigate the effects of different organic waste-derived feeds on the growth, Feed Conversion Ratio (FCR), bioconversion, survival, and nutritional composition, with a focus on Total Polyphenol Concentration (TPC) of black soldier fly larvae. By examining how various waste substrates influence the nutrient profile of the larvae with an emphasis on polyphenols, this research aims to provide insights into optimizing BSFL as a functional additive in aquaculture diets, contributing to both waste valorization and sustainable feed production.
2. Materials and Methods
2.1. Waste Collection and Preparation
Five feeds were tested which were comprised of organic waste from different commercial sectors. Kitchen Waste (KW, 50/50 lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potato, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO). KW was sourced from Salt Waterfront Restaurant, AS was sourced from J&C Tropicals, and AS and AO were sourced from Atlantic Sapphire; all waste sources were in the locality of Miami, Florida. KW, AW, and AO were chopped into smaller pieces; all samples were kept frozen until larval feeding with a 12hr thawing period before each feeding.
2.2. Larval Rearing and Experimental Design
The BSFL were reared in an enclosure where temperatures was 27.1 °C ± 0.2, humidity 76.4% ± 0.3, and D:L was 24:0. The larvae (5-day old) were soured from Symtom Black Soldier Fly (College Station, Texas) and placed in the experimental containers with their respective feed. Containers had a tight-fitting lid and 10 small holes with a 500μm screen to prevent escapes and allow for air circulation. There were five dietary treatments with four replicates each. Each replicate consisted of a container that received 200 larvae, 60g of Hydrated Coconut Coir (HCC), and a feed amount equal to 500% the larval biomass of each respective container. Quantification of biomass, feeding, and replacement of HCC occurred on Day 0 and every 3 days until Day 15 which is when the experiment was concluded. Feeding and substrate replacement were not done on Day 15. To separate larvae and HCC, a 500μm screen was used as a sieve. Initial and final biomass were used to calculate growth and biomass gain; initial and final number of larvae were used to calculate survival; biomass gain and feed given to each container were used to calculate FCR and bioconversion. The FCR value in this manuscript is not an accurate FCR quantifying the actual feed consumed as leftover feed was homogenously mixed with the HCC; excess feed and HCC were removed and disposed. The FCR presented here uses the amount of feed added to each container instead of the total feed consumed.
2.3. Analytical Analysis
2.3.1. Biomass Gain, Survival, Feed Conversion Ratio, Bioconversion, and Protein Efficiency Ratio
The effect of feed on larval performance, FCR, bioconversion, and nutritional composition were evaluated. The following are calculations for biomass gain, survival, FCR, and bioconversion:

2.3.2. Nutritional Composition
Larvae from each treatment were pooled together, dried at 105 °C for 24hrs, and vacuum packed for analysis. Fresh feeds were dried at 105 °C for 24hrs, and vacuum packed for analysis. Larvae and feeds were sent to an external NATA accredited laboratory for nutrient analysis.
Protein was quantified by combusting the sample in pure oxygen at 950°C, converting nitrogen oxides to nitrogen, and measuring nitrogen via thermal conductivity after removing water, oxygen, and carbon dioxide. The nitrogen content was multiplied by 6.25 to estimate protein content [15]. Fat content was determined by the Folch technique, involving chloroform/methanol (2:1) extraction, with BHT solution added to prevent oxidation. The extract was further processed with a sodium chloride/methanol solution, and the solvent removed by rotary evaporation (in vacuo) for gravimetric determination of lipid content [16]. Energy was determined by isoperibolic bomb calorimetry in a Parr 6200 oxygen bomb calorimeter (Par Instrument Company, Moline, IL, USA) [17]. Amino acids were quantified by hydrolyzing the samples in 6M HCl followed by Liquid Chromatography-Photodiode-Array-Mass Spectrometry (LC-PDA-MS) analysis, in accordance with NMI in-house method VL 450.The fatty acid profile is determined by gas chromatography on the hydrolyzed and esterified methyl esters [18,19].
2.3.3. Total Polyphenol Concentration
Polyphenol concentration of BSFL was evaluated through the Folin & Ciocalteu’s method [20]. Briefly, 0.5 mL of sample extract was mixed with 2.5 mL of Folin-Ciocalteu reagent (MERK, previously diluted 1:10 with distilled water) in a test tube. After standing for 5 minutes, 2 mL of 7.5% sodium carbonate solution was added, and the mixture was incubated in the dark at room temperature for 30 minutes to allow color development. The absorbance of the resulting blue complex was measured at 765 nm using a UV-Vis spectrophotometer. Total polyphenol content was calculated based on a standard curve generated with gallic acid and expressed as mg of gallic acid equivalents (GAE) per gram of sample. All measurements were performed in quadruplets to ensure accuracy.
2.4. Total Polyphenol Concentration
The statistical analysis was performed using GraphPad Prism 8 (v 8.0.1). The results of the experiment were subjected to one-way Analysis of Variance (ANOVA) to evaluate and compare the effect of the different feeds on BSFL performance. ANOVA was followed by Tukey's Multiple Comparison Test as a post hoc analysis to assess statistical differences between the means of the different groups, with a significance level set at 5% (p < 0.05). The data is presented in mean ± standard error of the mean (SEM).
3. Results
3.1. Growth and Survival
Figure 1 shows the growth performance in terms of container biomass affected by the different feed treatments, with sampling intervals on Day 0, 3, 6, 9, 12, and 15. The initial mean container biomass was 2.34g ± 0.03 with an average larval weight of 12mg ± 0.1. All feed treatments promoted larval growth. The feed treatment that had the highest final biomass was MX (34.8g ± 1.7), followed by AO (32.3g ± 0.6), AS (19.2g ± 0.3), AW (9.0g ± 0.2), and KW (7.0g ± 0.1). No significant differences (p > 0.05) were found between MX and AO; and AW and KW; both groups were significantly different (p < 0.05) to each other and to AS. The biomass of the AO treatment began an asymptotic phase around Day 9. Table 1 shows that over 90% of the larvae from all treatments survived, with no significant differences (p > 0.05) between groups. The highest survival was achieved with KW (98.9% ± 0.6), followed by MX (98.3% ± 1.2), AS (97.6% ± 0.3), AO (95.0% ± 1.51), and AW (93.9% ± 2.0).
3.2. Feed Conversion Ratio, Bioconversion, and Protein Efficiency Ratio
Table 1 shows FCR, bioconversion, and PER. Feed type had a significant effect on larval efficiency and bioconversion. The lowest FCR was achieved with MX (15.2 ± 0.3) which was statistically different (p < 0.05) from all other treatments. The FCR of MX was followed by AS (17.5 ± 0.2), AO (19.4 ± 0.1), AW (27.4 ± 0.7), and KW (33.5 ± 0.9). There was no significant difference (p > 0.05) between AO and AS. In terms of bioconversion there was significant differences (p < 0.05) between all treatments. The highest bioconversion was achieved by MX (7.1% ± 0.2), followed by AS (6.5% ± 0.1), AO (5.6% ± 0.1), AW (4.9% ± 0.1), and KW (4.6% ± 0.1). In terms of PER, the highest value was achieved by AW (69.5% ± 1.7) which was statistically different (p < 0.05) from all other treatments. The PER of AW was followed by MX (29.1% ± 0.6), KW (21.2% ± 0.6), AS (19.3% ± 0.2), and AO (13.0% ± 0.1). However, there was no significant difference between KW and AS (p > 0.05).

Table 1.
Black Soldier Fly Larvae rearing parameters when fed the different feed treatments.
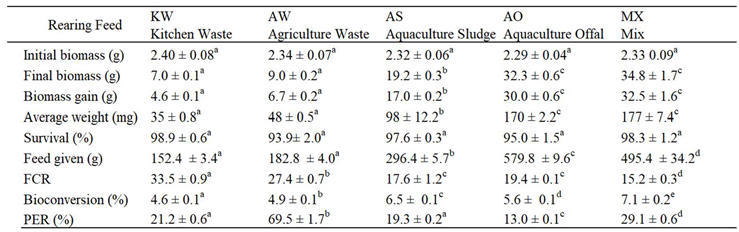
Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potatoes, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO). Data presented is mean ± SEM for quadruplets, means denoted by a different letter indicate significant difference between treatments (p < 0.05).
3.3. Nutritional Composition
3.3.1. Feeds
Nutrients of the different BSFL feeds differed based on their respective origin (Table 2). Crude protein ranged from 39.9% in AO to 5.3% in AW. Crude lipid ranged from 60.4% in AO to 2.5% in AW. Gross energy ranged from 26.5% in MX to 17.5% in AW and AS. Ash ranged from 29.0% in AS to 2.4% in AO. Total Amino Acid (TAA) ranged from 50.4% in AO to 4.6% in AW. The sum of Indispensable Amino Acids (IAA) ranged from 16.3% in AO to 1.3% in AW; and Dispensable Amino Acids (DAA) ranged from 17.9% in AO to 1.9% in AW. Saturated Fatty Acids Capric (C10:0) and Lauric (C12:0) were negligible in all larval feed. Omega-3 ranged from 6.6% in AO to 0.4% in KW. EPA ranged from 1.6% in AS to <0.1% in KW. DHA ranged from 2.8% in AO to <0.1% in KW and AW.
3.3.2. Larvae
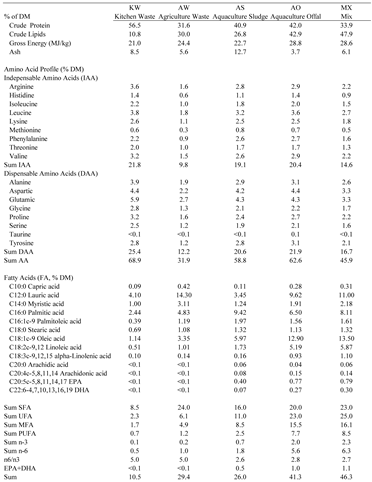
Nutritional composition of BSFL fed their feed treatments is presented in Table 3. Crude protein ranged from 56.5% in KW to 31.6% in AW. Crude lipid ranged from 47.9% in MX to 10.8% in KW. Gross energy ranged from 28.8% in AO to 21.0% in KW. Ash ranged from 12.7% in AS to 3.7% in AO. TAA ranged from 68.9% in KW to 31.9% in AW. The sum of IAA ranged from 21.8% in KW to 9.8% in AW; and DAA ranged from 25.4% in KW to 12.2% in AW. Saturated Fatty Acids Capric (C10:0) ranged from 0.4% in AO to 0.1% in KW; Lauric (C12:0) ranged from 14.3% in AW to 3.4% in AS. Omega-3 ranged from 2.3% in MX to 0.1 in KW. EPA ranged from 0.8% in MX to <0.1 in KW and AW. DHA ranged from 0.3% in MX to <0.1 in KW and AW.
3.3.3. Total Polyphenol Concentration
TPC of the different BSFL varied significantly depending on the type of waste the larvae consumed and is presented in Figure 2. The treatments that exhibited the highest TPC were MX (7.4mg GAE/100g ± 1.0), followed by KW (7.2mg GAE/100g ± 1.9), and AW (5.8mg GAE/100g ± 0.9), without significant different to each other (p > 0.05). AS (0.9mg GAE/100g ± 0.4) and AO (0.6mg GAE/100g ± 0.3) had low TPC and were not statistically different from each other (p > 0.05). MX, KW and AS were statistically different to AS and AO (p < 0.05).
4. Discussion
This study evaluated the effects of different organic waste-derived feeds on the growth, survival, feed conversion ratio, and bioconversion of Hermetia illucens larvae over a 15-day period. The feed types—Kitchen Waste (KW), Agricultural Waste (AW), Aquaculture Sludge (AS), Aquaculture Offal (AO), and Mix (MX)—varied in nutrient composition, which significantly influenced the larvae’s performance across all parameters. The findings highlight the potential of H. illucens as efficient bio-converters of waste into valuable biomass, with diet composition playing a critical role in optimizing larval growth, waste conversion, and nutrient accumulation.
The larvae fed on AO and MX exhibited the highest larval mean weight, with MX reaching 177mg and AO 170mg. AO was comprised of salmon offal which contains high levels of proteins and lipids, and was a homogenous mix of different organs, while MX was a homogonous mix of plant and animal-based feed. The results of biomass gain and growth in this research are consistent with prior research indicating that high-protein and fat-rich diets as well as balanced nutrient profiles promote enhanced larval growth, biomass accumulation, and are essential for larval tissue development [21,22,23,24]. In contrast, larvae fed plant-based diets like KW and AW exhibited significantly lower larval mean weight (0.035g and 0.048g, respectively), possibly due to the feed’s lack of nutrients and susceptibility to microbial spoilage due its high moisture content, which could have reduced nutrient availability and digestibility [24,25].
Feed conversion ratio, measured as the ability of a unit of feed to produce a unit of larval biomass, was lowest with MX, AS and AO. This reflects the larval superior ability in converting nutrient-dense, high-protein, and fat-rich diets into biomass. This finding is consistent with studies showing that nutrient-dense feeds, particularly those rich in protein and fat, promote more efficient feed conversion in BSFL [26]. Similarly, bioconversion, defined as the ability of larvae to convert organic waste into biomass, was highest in MX and AS. The high bioconversion rates observed reinforce BSFL’s capacity as efficient upcyclers of organic waste into valuable biomass, a finding that mirrors prior research assessing their bioconversion capabilities on various waste substrates [7]. In comparison, larvae fed plant-based diets like KW and AW exhibited the highest feed conversion ratio and lowest bioconversion, due to the lack of proteins and lipids and high moisture content which could dilute the nutrient density as was reported in previous studies [27]. Although these diets did not perform as well as the animal-based feeds, they still resulted in notable biomass production, highlighting the versatility of BSFL in processing various waste streams. This versatility is critical in waste valorization strategies as BSFL can successfully convert lower-quality waste into usable biomass.
Survival rates were consistently high across all dietary treatments, ranging from 93% to 99%, with no statistically significant differences between the groups. This suggests that despite variations in nutrient composition among the different feeds, Hermetia illucens larvae demonstrated resilience and adaptability to a wide range of substrates. This finding is in line with previous research showing that BSFL maintain high survival rates across various substrates, provided conditions are not overly detrimental [27,24,28]. The ability to maintain high survival rates across different feed types, including both nutrient-rich and nutrient-poor feed sources, highlights the suitability of BSFL for diverse waste management applications. It is important to mention that any negative compounding effects that the dietary treatments might have on the larvae and nutrient uptake were negated due to leftover feed and HCC being replaced with a new batch every 3 days. Excess moisture from KW and AW, excess leached oils from AO, and excess salt from AS were regularly removed from the culture environment. It is important to note that this research was carried out at an experimental scale, and transition to a larger commercial scale operation will warrant more understanding of the effects of the different diets on larval development and nutrient accumulation.
BSFL require certain key nutrients to grow through the different developmental stages to reach pupation. Environmental parameters and factors such as density will also influence larval development and growth. Not only do nutrients need to be present in larval feed for optimal development, but those nutrients need to be bioavailable. This nutrient bioavailability is dependent on the organism consuming that nutrient and its developmental stage, and the feed type being provided [24]. The nutrient profiles of black soldier fly larvae (BSFL) fed different feeds exhibit substantial variation in protein, lipid, amino acid, and fatty acid content, reflecting the significant influence of feed composition on larvae nutritional quality. KW larvae obtained the highest protein level (56.5%) among the experimental treatments, this in line with findings that vegetable waste can support high protein synthesis in BSFL [29]. This is an interesting observation as KW feed contained one of the lowest crude protein levels (14.1%), but the KW larvae accumulated the highest crude protein of all treatments (56.5%). A similar pattern occurred with AW feed that had a protein level of 5.3% which resulted in AW larvae with a protein level of 31.6%. This could indicate that BSFL are more efficient in converting vegetable rather than animal proteins. This is further validated by looking at the PER, where larvae fed with plant-based feeds (AW, MX, and KW) had a higher ratio value than larvae fed with animal-based feed (AS and AO). This protein accumulation pattern was similarly observed by Lalander et al. (2019) [24]. The authors saw that low-density feeds like fruits, vegetables and sludge allowed for a higher protein accumulation rate than high-density feeds like slaughter waste which impaired protein accumulation.
The MX larvae treatment demonstrated the highest lipid content (47.9%) followed by AO (42.9%), which correlates with previous research indicating that protein- and fat-rich substrates significantly increase larvae lipid accumulation [30]. The AW feed, despite having the lowest crude lipid (2.5%), produced a larva that contained average crude lipid levels (30.0%) when compared to the other treatments. This could be attributed to the carbohydrates present in AW feed that are bio-converted into lipid [31]. A noteworthy remark is that even though the AO larvae were fed with a feed source with 60.4% lipid, the larvae accumulating only 42.9% lipid, which might indicate that a lipid accumulation of more than ~40% is not physiologically possible. The ash content of the BSFL reared in this experiment was directly affected by the ash content of their respective feed, a similar result obtained by Spranghers et al. (2017) who saw a direct proportionality of ash in the larvae to the feed [29].
This research demonstrates that when provided with plant-based feeds, the larvae are better able to accumulate amino acids relative to the feed. The amino acid accumulation rate observed here is similar to that of Lalander et al. (2019) who saw that simple and low protein feeds allowed the larvae to accumulate higher rates of protein when compared to complex and protein dense feeds [24]. This indicates that plant-based feeds, even though lacking high levels of amino acids, allow the larvae to accumulate those same nutrients at levels significantly higher than their feed. Interestingly, even though taurine was quantified in the larval tissue in small amounts, it failed to properly accumulate more than any of the feeds. This could possibly indicate a physiological lack of appropriate metabolic pathways for taurine deposition. Gianetto et al. (2020) demonstrated that BSFL have a series of genes that regulate taurine biosynthesis, and when fed with a vegetable-based feed (40% pear, 45% banana, 5% tomatoes, 10% various leafy green vegetables) the prepupae contained 45 ± 0.01 mg/kg of taurine (0.0045%) [32]. The study demonstrated that the gene expression was developmentally regulated, with a two-fold increase in expression for prepupa than larvae. This could suggest that taurine biosynthesis has an active role during late-developmental stages. The authors did not provide the concentration of taurine in the larval feed, and so it is not fully possible to determine whether the prepupae accumulated taurine at a higher concentration than the feed. This might indicate that BSFL can synthesize taurine de novo but are unable to efficiently accumulate the amino acids in its tissue. Similarly to Eggink et al. (2022), in this research the taurine larval levels were the lowest amongst all amino acids, but its concentration was higher in larvae fed marine-based feed (AS, AO, and MX) rather than vegetable-based (KW and AW). Considering that both the larvae in this research and those of Eggink et al. (2022) were not starved prior to harvest, the higher taurine levels could be related to leftover feed in the intestinal tract (gut loading) instead of de novo synthesis [33].
Larvae typically exhibit high levels of saturated fatty acids (SFAs), with lauric acid (C12:0) being particularly abundant, followed by palmitic acid (C16:0) and oleic acid (C18:1). This is especially true when larvae are reared on substrates rich in SFAs. For instance, larvae fed on coconut oil-enriched diets show increased medium-chain lauric acid content; larvae fed on olive-oil enriched diets show increased monounsaturated oleic acid content; and larvae fed on soybean oil-enriched diets show increased polyunsaturated linoleic acid content [34]. BSFL can synthesize and accumulate a variety of SFA such as capric, lauric and myristic [35]. This is confirmed by our results as these fatty acids were present in the larvae yet almost absent in the experimental feeds, indicating a level of synthesis and deposition. In terms of long-chain polyunsaturated fatty acids (LC-PUFA), BSFL have a limited capacity to synthesize these fatty acids but can deposit them with certain limitations [36]. In the case of AO and MX, the levels of ALA, EPA, and DHA in the larvae were lower than those of their respective feed but higher than then other dietary treatments. This is in line with St. Hilaire et al. (2007) who saw an increase in larval omega-3 fatty acids when fed with fish offal (homogenized heads, viscera, and some bony structures from rainbow trout), and as well as Erbland et al. (2020) who fed graded levels of salmon oil [30,37]. Interestingly, MX was comprised of 50% salmon offal and AO was comprised of 100% salmon offal, yet the larval levels of these long-chain PUFA was almost the same, which seems to indicate that BSFL cannot deposit these fatty acids past a certain point and become saturated.
This plateau effect may be due to metabolic constraints or saturation of lipid storage mechanisms within the larvae [34]. In fact, except for capric, lauric and myristic acid in the AO larvae, all fatty acids were lower in the larvae than in the feed, possibly indicating a fatty acid dietary saturation that limits fatty acid deposition.
The higher polyphenol accumulation observed in BSFL fed KW, AW, and MX can be linked to the naturally high TPC in fruits and vegetables. Depending on the different cultivars, origin of culture, culture conditions, organic/non, fruits and vegetables can have differing biochemical profiles [38]. Tomatoes, sweet potatoes, lettuce, dragon fruit, taro, and yuca (used in this research) have been observed to have a TPC of 250, 150, 100, 40, 25, 10 mg GAE/100g FW, respectively [39,40,41,42,43,44,45]. These concentrations reflect the natural antioxidant capacity of these foods, which seem to influence the polyphenol accumulation in larvae when used as feed. This variation helps explain why larvae fed on plant-based wastes exhibit higher TPC compared to those fed on animal-based wastes like aquaculture offal. However, the mix of plant-based and animal-based waste (MX treatment), despite containing 50% of low-polyphenol ingredients like aquaculture offal and sludge, was still able to yield larvae with the highest polyphenol concentrations (although not statistically different from KW and AW). A possible hypothesis is that the larvae fed KW and AW reached the maximum upper limit of polyphenols accumulation in their tissue, and a higher concentration of these compounds was not physiologically possible. This similar limit can be observed through bioenergetic efficiency in fish, where beyond a certain dietary protein level, nitrogenous waste production increases due to unused amino acids being broken down and not fully utilized [46]. On the other hand, AS and AO did not support polyphenol accumulation as these substrates are largely devoid of plant-based antioxidants. This is consistent with research showing that polyphenols are predominantly found in plant materials, and that BSFL’s ability to bioaccumulate polyphenols is dependent on the presence of these compounds in their feed [47,48].
It must be noted that the evaluated feeds might not be directly linked to the different nutritional profiles of the larvae, but these differences might be coming from a difference in larval developmental stages. If nutrients are limited or scarce, the larvae can extend their larval stage to increase their time to accumulate the necessary nutrients to transition to the next life stage [25,49]. Additionally, the nutritional profile of the larvae changes throughout the life stages of the larvae, as demonstrated by Liu et al. (2017) [40]. In the presented manuscript, all larvae from all treatments were harvested on Day 15 irrespective of developmental stage. The hypothesis that the larvae of the different treatments were at differing stages was supported by the coloration of the different treatments, with AO and MX being a darker color than KW and AW, indicating being closer to pupation.
5. Conclusions
This study highlights the potential of Black Soldier Fly Larvae (BSFL) as a sustainable solution for waste management and aquafeed production. BSFL efficiently convert organic waste into nutrient-rich biomass, supporting circular economy principles by reducing waste and optimizing resources. Feed composition significantly influenced larval growth, feed efficiency, and polyphenol content, with plant-based feeds showing higher Total Polyphenol Concentrations (TPC) than animal-based feeds. Mix, Kitchen Waste, and Agricultural Waste recorded TPCs of 7.4, 7.2, and 5.8 mg GAE/100g, while Aquaculture Sludge and Offal had 0.9 and 0.6 mg GAE/100g. These findings advance sustainable aquaculture by reducing reliance on traditional protein sources and promoting fish health through a functional feed ingredient, aligning with global sustainability goals.
Author Contributions
Conceptualization, Julio Camperio, Justin Simonton and Eli Paresky; Data curation, Julio Camperio, Jorge Parodi and Jorge Suarez; Formal analysis, Julio Camperio, Jorge Parodi and Jorge Suarez; Funding acquisition, Julio Camperio, Eli Paresky, Jorge Suarez and Daniel Benetti; Investigation, Julio Camperio, Justin Simonton, Eli Paresky, Jorge Parodi and Jorge Suarez; Methodology, Julio Camperio, Justin Simonton, Eli Paresky, Jorge Parodi and Jorge Suarez; Project administration, Julio Camperio, Jorge Suarez and Daniel Benetti; Resources, Julio Camperio and Jorge Suarez; Supervision, Jorge Suarez and Daniel Benetti; Validation, Julio Camperio, Justin Simonton, Eli Paresky, Jorge Parodi and Jorge Suarez; Visualization, Julio Camperio, Justin Simonton, Eli Paresky, Jorge Parodi and Jorge Suarez; Writing – original draft, Julio Camperio; Writing – review & editing, Justin Simonton, Eli Paresky, Jorge Parodi, Jorge Suarez and Daniel Benetti.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
This project was made possible thanks to entities that donated their organic waste: Salt Waterfront Restaurant, J&C Tropicals, and Atlantic Sapphire. Additionally Carlos Tudela for assisting in sampling.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| BSFL | Black Soldier Fly Larvae |
| KW | Kitchen Waste |
| AW | Agriculture Waste |
| AS | Aquaculture Sludge |
| AO | Aquaculture Offal |
| MX | Mix |
| FCR | Feed Conversion Ratio |
| PER | Protein Efficiency Ratio |
| GAE | Gallic Acid Equivalent |
| BSFLM | Black Soldier Fly Larvae Meal |
| HCC | Hydrate Coconut Coir |
| TAA | Total Amino Acids |
| IAA | Indispensable Amino Acids |
| DAA | Dispensable Amino Acids |
| LC-PUFA | Long Chain – Polyunsaturated Fatty Acids |
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Turchini, G.M.; Trushenski, J.T.; Glencross, B.D. Thoughts for the future of aquaculture nutrition: Realigning perspectives to reflect contemporary issues related to judicious use of marine resources in aquafeeds. North Am. J. Aquac. 2019, 81, 13–39. [Google Scholar] [CrossRef]
- Booman, M.; Forster, I.; Vederas, J.C.; Groman, D.B.; Jones, S.R.M. Soybean meal-induced enteritis in Atlantic salmon (Salmo salar) and Chinook salmon (Oncorhynchus tshawytscha) but not in pink salmon (O. gorbuscha). Aquaculture 2018, 483, 238–243. [Google Scholar] [CrossRef]
- van Huis, A. Insects as food and feed, a new emerging agricultural sector: A review. J. Insects Food Feed 2019, 6, 27–44. [Google Scholar] [CrossRef]
- Schiavone, A.; De Marco, M.; Martínez, S.; Dabbou, S.; Renna, M.; Madrid, J.; Gasco, L. Nutritional value of a partially defatted and a highly defatted black soldier fly larvae meal for broiler chickens: Apparent nutrient digestibility, apparent metabolizable energy, and apparent ileal amino acid digestibility. J. Anim. Sci. Biotechnol. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Heuze, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Gold, M.; Tomberlin, J.K.; Diener, S.; Zurbrügg, C.; Mathys, A. Decomposition of biowaste macronutrients, microbes, and chemical properties during the conversion process by black soldier fly larvae (Hermetia illucens). Sci. Total Environ. 2018, 613, 130–141. [Google Scholar] [CrossRef]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. 2016, 100, 653–664. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Parodi, A.; Leip, A.; De Boer, I.J.M.; Slegers, P.M.; Ziegler, F.; Temme, E.H.M.; van Zanten, H.H.E. The potential of future foods for sustainable and healthy diets. Nat. Sustain. 2018, 1(12), 782–789. [Google Scholar] [CrossRef]
- Lock, E.R.; Arsiwalla, T.; Waagbø, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postsmolt. Aquac. Nutr. 2021, 27(2), 568–579. [Google Scholar] [CrossRef]
- Moutinho, S.; Oliva-Teles, A.; Fontinha, F.; Martins, N.; Monroig, Ó.; Peres, H. Black soldier fly larvae meal as a potential modulator of immune, inflammatory, and antioxidant status in gilthead seabream juveniles. Comp. Biochem. Physiol. B 2024, 271, 110951. [Google Scholar] [CrossRef] [PubMed]
- Weththasinghe, P.; Rocha, S.D.C.; Øyås, O.; et al. Modulation of Atlantic salmon (Salmo salar) gut microbiota composition and predicted metabolic capacity by feeding diets with processed black soldier fly (Hermetia illucens) larvae meals and fractions. Anim. Microbiome 2022, 4, 9. [Google Scholar] [CrossRef]
- AOAC Official Method 990.03. Protein (crude) in animal feed, combustion method. Official Methods of Analysis of AOAC International, 2006, 18th Ed.; AOAC International: Gaithersburg, MD, USA.
- Bligh, E.G.; Dyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- International Organization of Standardization. Animal feeding stuffs, animal products, and faeces or urine — Determination of gross calorific value — Bomb calorimeter method, ISO Standard No. 9831.1998. ISO 1998.
- AOAC Official Method 969.33. Fatty acids in oils and fats. Official Methods of Analysis of AOAC International, 1969, AOAC International: Gaithersburg, MD, USA.
- AOAC Official Method 969.22. Methyl esters of fatty acids in oils and fats. Gas chromatographic method. Official Methods of Analysis of AOAC International, 1997, AOAC International: Gaithersburg, MD, USA.
- Olivares-Ferretti, P.; Chavez, V.; Hernandez, K.; Parodi, J. Polyphenols extracts from Didymosphenia geminata (Lyngbye) Schmidt altered the motility and viability of Daphnia magna. Res. Square 2021. [CrossRef]
- Belperio, S.; Cattaneo, A.; Nannoni, E.; Sardi, L.; Martelli, G.; Dabbou, S.; Meneguz, M. Assessing substrate utilization and bioconversion efficiency of black soldier fly (Hermetia illucens) larvae: Effect of diet composition on growth and development temperature. Animals 2024, 14(9). [CrossRef]
- Chia, S.Y.; Tanga, C.M.; Khamis, F.M.; Mohamed, S.A.; Salifu, D.; Sevgan, S.; Ekesi, S. Threshold temperatures and thermal requirements of black soldier fly Hermetia illucens: Implications for mass production. PLoS One 2020, 15(1), e0228541. [Google Scholar] [CrossRef]
- Nyakeri, E.M.; Ogola, H.J.O.; Ayieko, M.A.; Amimo, F.A. An open system for farming black soldier fly larvae as a source of proteins for small-scale poultry and fish production. J. Insects Food Feed 2017, 3(1), 51–56. [Google Scholar] [CrossRef]
- Lalander, C.; Diener, S.; Zurbrügg, C.; Vinnerås, B. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). J. Clean. Prod. 2018, 233, 116–126. [Google Scholar] [CrossRef]
- Bekker, N.S.; Heidelbach, S.; Vastergaard, S.Z.; Nielsen, M.E.; Riisgaard-Jensen, M.; Zeuner, E.J.; Bahrndorff, S.; Eriksen, T. Impact of substrate moisture content on growth and metabolic performance of black soldier fly larvae. Waste Manag. 2021, 127, 73–79. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166(9), 761–770. [Google Scholar] [CrossRef] [PubMed]
- Diener, S.; Zurbrügg, C.; Gutiérrez, F.R.; Nguyen, D.H.; Morel, A.; Koottatep, T.; Tockner, K. Black soldier fly larvae for organic waste treatment—Prospects and constraints. Proceedings of WasteSafe 2011 – 2nd International Conference on Solid Waste Management in the Developing Countries, Khulna, Bangladesh, 13-15 February 2011. [Google Scholar]
- Li, R.; Lin, T.; Fan, X.; Dai, X.F.; Huang, J.H.; Zhang, Y.F.; Guo, R.B.; Fu, S.F. Effects of salinity in food waste on the growth of black soldier fly larvae and global warming potential analysis. Chem. Eng. J. 2024, 480. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97(8), 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aquac. Soc. 2007, 38(2). [CrossRef]
- Liu, Y.; Liu, J.; He, J.; Lu, H.; Sun, S.; Ji, F.; Dong, X.; Bao, Y.; Xu, J.; He, G.; Xu, W. Chronological and carbohydrate-dependent transformation of fatty acids in the larvae of black soldier fly following food waste treatment. Molecules 2023, 28, 41903. [Google Scholar] [CrossRef]
- Giannetto, A.; Oliva, S.; Riolo, K.; Savastano, D.; Parrino, V.; Cappello, T.; Maisano, M.; Fasulo, S.; Mauceri, A. Waste valorization via Hermetia illucens to produce protein-rich biomass for feed: Insight into the critical nutrient taurine. Animals 2020, 10(9), 1710. [Google Scholar] [CrossRef]
- Eggink, K.M.; Lund, I.; Pedersen, P.B.; Hansen, B.W.; Dalsgaard, J. Biowaste and by-products as rearing substrates for black soldier fly (Hermetia illucens) larvae: Effects on larval body composition and performance. PLoS One 2022, 17(9), e0275213. [Google Scholar] [CrossRef]
- Li, X.; Dong, Y.; Sun, Q.; Tan, X.; You, C.; Huang, Y.; Zhou, M. Growth and fatty acid composition of black soldier fly (Hermetia illucens) larvae are influenced by dietary fat sources and levels. Animals 2022, 12(4), 486. [Google Scholar] [CrossRef]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens) – Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef]
- Barroso, F.G.; Sanchez-Muros, M.J.; Rincon, M.A.; Rodriguez-Rodriguez, M.; Fabrikov, D.; Morote, E.; Guil-Guerrero, J.S. Production of n-3-rich insects by bioaccumulation of fishery waste. J. Food Compos. Anal. 2019, 82, 103237. [Google Scholar] [CrossRef]
- Erbland, P.; Alyokhin, A.; Perkins, B.; Peterson, M. Dose-dependent retention of omega-3 fatty acids by black soldier fly larvae (Diptera: Stratiomyidae). J. Econ. Entomol. 2020, 113(3), 1221–1226. [Google Scholar] [CrossRef]
- Tylewicz, U.; Nowacka, M.; Marin-Garcia, B.; Wiktor, A. Target sources of polyphenols in different food products and their processing by-products. In: Polyphenols: Properties, Recovery, and Applications; Woodhead Publishing, 2018, pp. 135-175. [CrossRef]
- García-Valverde, V.; Inmaculada, N.G.; García-Alonso, J.; Periago, M.J. Antioxidant bioactive compounds in selected industrial processing and fresh consumption tomato cultivars. Food Bioprocess Technol. 2013, 6, 391–402. [Google Scholar] [CrossRef]
- Islam, M.S.; Yoshimoto, M.; Yahara, S.; Okuno, S.; Ishiguro, K.; Yamakawa, O. Identification and characterization of foliar polyphenolic composition in sweet potato (Ipomoea batatas L.) genotypes. J. Agric. Food Chem. 2002, 50(13), 3718-3722. [CrossRef]
- Sun, Y.; Pan, Z.; Yang, C.; Jia, Z.; Guo, X. Comparative assessment of phenolic profiles, cellular antioxidant and antiproliferative activities in ten varieties of sweet potato (Ipomoea batatas) storage roots. Molecules 2019, 24(24), 4476. [Google Scholar] [CrossRef] [PubMed]
- Vella, F.; Calandrelli, R.; Cautela, D.; Laratta, B. Screening of bioactivity in extracts from different varieties of lettuce. In Proceedings of the International Electronic Conference on Plant Science; 2020. [Google Scholar] [CrossRef]
- Brar, J.S.; Sharma, S.; Kaur, H.; Singh, H.; Naik, E.K.; Adhikary, T. Phytochemical properties, antioxidant potential and fatty acids profiling of three dragon fruit species grown under sub-tropical climate. Not. Bot. Horti Agrobot. Cluj-Napoca 2023, 51(3). [CrossRef]
- Ouédraogo, N.; Sombie, P.A.E.D.; Traore, R.E.; Sama, H.; Bationo/Kando, P.; Sawadogo, M.; Lebot, V. Nutritional and phytochemical characterization of taro (Colocasia esculenta (L.) Schott) germplasm from Burkina Faso. J. Plant Breed. Crop Sci. 2023, 15(1), 32-41. [CrossRef]
- Faezah, N.; Aishah, S.; Kalsom, U. Comparative evaluation of organic and inorganic fertilizers on total phenolic, total flavonoid, antioxidant activity, and cyanogenic glycosides in cassava (Manihot esculenta). Afr. J. Biotechnol. 2013, 12(18), 2414–2421. [Google Scholar] [CrossRef]
- Bureau, D.P.; Kaushik, S.J.; Cho, C.Y. Bioenergetics. In Fish Nutrition, 3rd ed.; Halver, J.E.; Hardy, R.W., Eds.; Academic Press, 2002, pp. 1-59.
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79(5), 727–747. [Google Scholar] [CrossRef]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81(1), 215S–217S. [Google Scholar] [CrossRef]
- Liu, Z.; Minor, M.; Morel, P.C.; Najar-Rodriguez, A.J. Bioconversion of three organic wastes by black soldier fly (Diptera: Stratiomyidae) larvae. Environ. Entomol. 2018, 47(6), 1609–1617. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, H.; Yang, Q.; Rehman, K.; Li, W.; Cai, M.; Li, Q.; Mazza, L.; Zhang, J.; Yu, Z.; Zheng, L. Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS One 2017, 12(8), e0182601. [Google Scholar] [CrossRef]
Figure 1.
Average container biomass during the experimental trial. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potato, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO). Data presented is mean ± SEM. Of quadruplets, means denoted by a different letter indicate significant differences between treatments (p < 0.05).
Figure 1.
Average container biomass during the experimental trial. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potato, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO). Data presented is mean ± SEM. Of quadruplets, means denoted by a different letter indicate significant differences between treatments (p < 0.05).
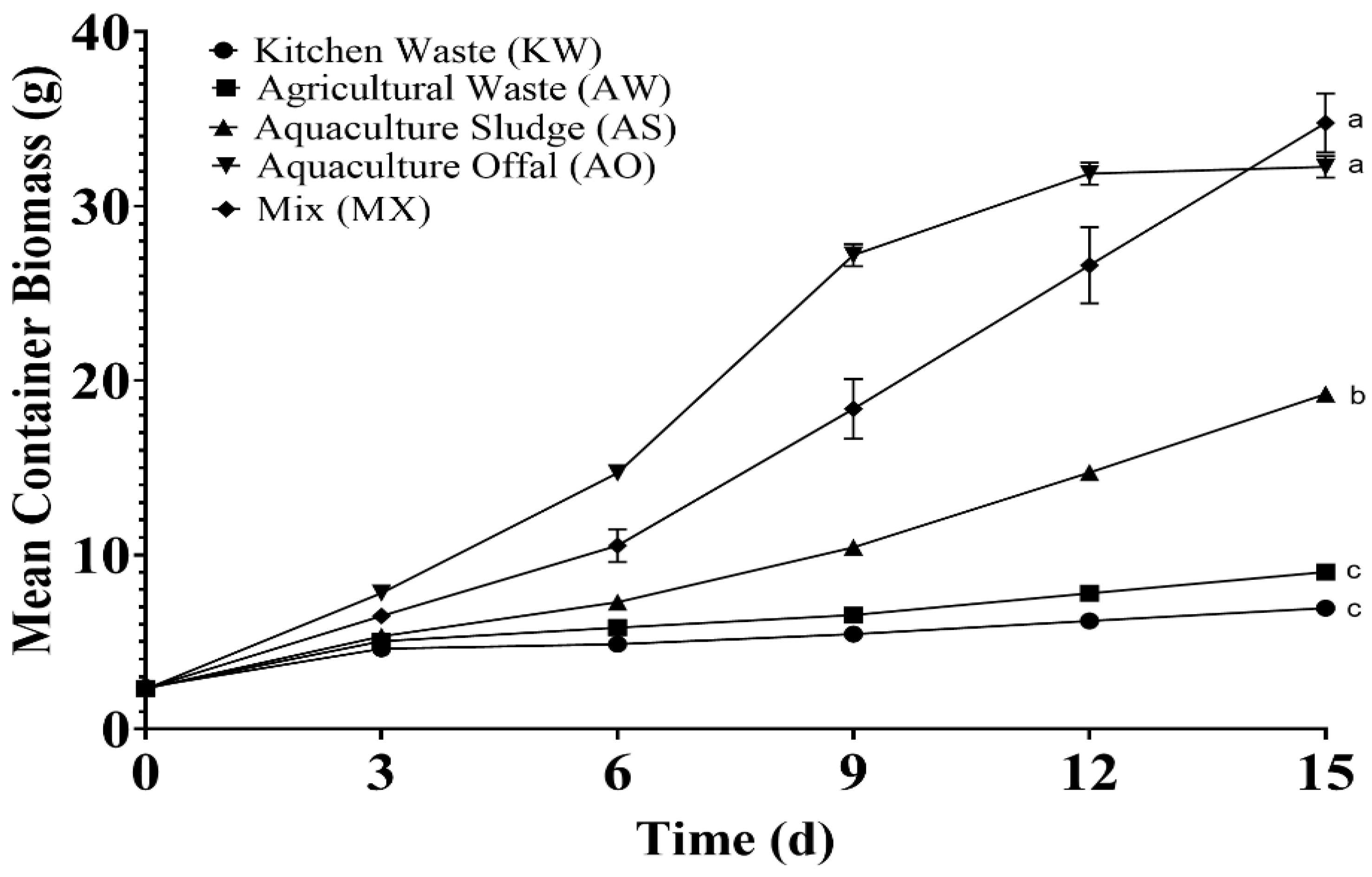
Figure 2.
Polyphenol concentration of the larvae fed the different dietary treatments. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potatoes, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO). Data presented is mean ± SEM for triplicates, means denoted by a different letter indicate significant difference between treatments (p < 0.05).
Figure 2.
Polyphenol concentration of the larvae fed the different dietary treatments. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potatoes, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO). Data presented is mean ± SEM for triplicates, means denoted by a different letter indicate significant difference between treatments (p < 0.05).
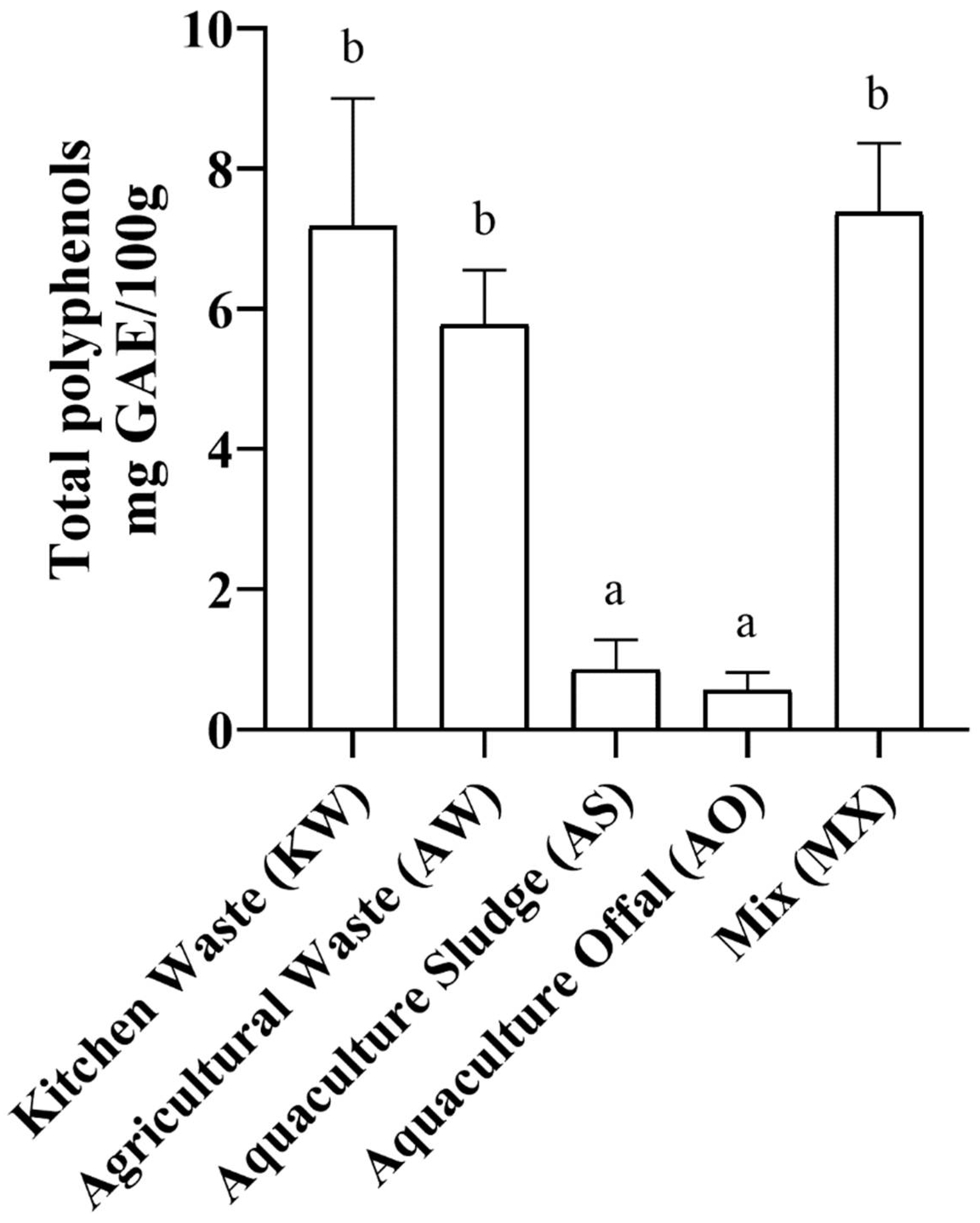
Table 2.
Proximate, amino acid, and fatty acid composition of the larval feed. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potatoes, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO).
Table 2.
Proximate, amino acid, and fatty acid composition of the larval feed. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potatoes, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO).
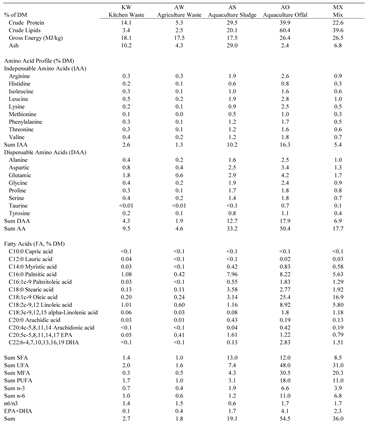
Table 3.
Proximate, amino acid, and fatty acid composition of the larval after consuming their respective feed for 15 days. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potatoes, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO).
Table 3.
Proximate, amino acid, and fatty acid composition of the larval after consuming their respective feed for 15 days. Kitchen Waste (KW, 50/50 romaine lettuce and tomatoes), Agricultural Waste (AW, 25/25/25/25 dragon fruit, sweet potatoes, yuca, and taro), Aquaculture Sludge (AS, waste sludge from a salmon RAS), Aquaculture Offal (AO, salmon offal post-processing), and Mix (MX, 25/25/25/25 of KW, AW, AS, and AO).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



