
Submitted:
03 December 2024
Posted:
04 December 2024
You are already at the latest version
Abstract
(1) Background: The RoXstaTM system has been developed as a potential diagnostic tool to profile the antioxidant activity of complex biological fluids. (2) Methods: This methodology was used to assess the ability of different biofluids (semen, blood plasma, serum, urine, saliva, follicular fluid and plant extracts) to suppress free radical formation, as well as scavenge a variety of toxic oxygen metabolites including free radicals and peroxides. (3) Results: Human semen was shown to have significantly (p < 0.001) more peroxide scavenging power than any other fluid tested, while urine was particularly effective in scavenging free radicals and preventing free radical formation. The powerful antioxidant properties of human semen were shown to reside within the seminal plasma (SP) fraction, rather than the spermatozoa, and to be resistant to snap freezing in liquid nitrogen. Moreover, comparative studies demonstrated that human SP exhibited significantly (p < 0.001) higher levels of antioxidant potential than any other species examined (stallion, bull, dog) and that this intense activity reflected the relative vulnerability of human spermatozoa to peroxide attack. (4) Conclusions: the RoXstaTM system provides valuable information on the antioxidant profile of complex biological fluids, supporting its diagnostic role in conditions associated with oxidative stress. Based on the results secured in this study, human semen is identified as a particularly rich source of antioxidants capable of scavenging both hydrogen-, and organic- peroxides, in keeping with the high susceptibility of human spermatozoa to peroxide-mediated damage.
Keywords:
antioxidant
; RoxStaTM system
; semen
; sperm motility
; blood
; urine
; saliva
; follicular fluid
; fruit juices
1. Introduction
A wide range of human and animal pathologies are thought to be related to the development of oxidative stress including cancer, neurodegenerative conditions, depression, multiple sclerosis, amyotrophic lateral sclerosis, cataracts, diabetes, cardiovascular disease and infertility [1,2]. Such associations have led to intense interest in the use of antioxidant supplements to restore the balance between reactive oxygen species (ROS) production and antioxidant protection. Unfortunately, the results of clinical trials addressing this topic have been variable and, ultimately, disappointing [3,4]. Apart from the pharmacological properties of the antioxidant agents themselves, one of the major issues with such evaluations relates to the selection of appropriate patients for treatment. Many clinical trials involving antioxidants have failed to select patients that are actually suffering from detectable oxidative stress and therefore might reasonably benefit from antioxidant supplementation. In the field of infertility research for example, males have been selected for such treatment because they exhibited oligozoospermia (low sperm numbers), asthenozoospermia (poor sperm motility) teratozoospermia (poor sperm morphology), high levels of DNA fragmentation, varicocele, prolonged idiopathic subfertility or recurrent pregnancy loss [4,5,6,7,8,9,10,11,12], rather than any direct evidence of heightened oxidative stress. Under these circumstances the results, not surprisingly, have been inconclusive because the etiologies underpinning these reproductive conditions are complex and, in some cases, may not involve any element of redox imbalance [13]. Furthermore, administering powerful antioxidants to patients who are not antioxidant deficient, brings with it a risk of creating reductive stress, which can be just as harmful as its oxidative counterpart [14].
With such poorly selected patient populations, it is entirely predictable that clinical trials addressing the value of antioxidants in the treatment of conditions such as male infertility will generate variable results; some patients will get better, some will get worse and, overall, any therapeutic benefit will just become lost in the statistical noise. On the rare occasion that such patients have actually been selected on the basis of verifiable oxidative stress, then the application of antioxidants has been found to generate positive clinical outcomes [15]. Thus, before we spend any more time and resource on clinical trials investigating the benefits of antioxidant therapy for infertility, or any other kind of pathology, we need to develop simple robust methodologies to test for the oxidative stress we are trying to counter [16].
To address this need, we have recently developed a novel suite of assays (the RoXsta™ system) to measure different aspects of antioxidant activity using ABTS (2,2’-azinobis-(3-ethylbenzthiazoline-6-sulphonic acid) as a redox sensor [17]. These assays were developed to measure the ability of biological fluids to suppress free radical formation as well as engage in the scavenging of free radicals, hydrogen peroxide and organic peroxides. Using human seminal fluid to evaluate the biological significance of these assays, we found that their combined output correlated well with such physiological parameters as sperm count, motility, the generation of ROS by the sperm mitochondria, sperm DNA damage and the presence of cytotoxic lipid aldehydes (specifically malondialdehyde) in semen [17]. In the present study we have examined the antioxidant profiles generated by these assays in a range of materials including blood plasma, serum, urine, saliva, follicular fluid and semen. The results confirm the capacity of these assays to provide rapid, effective assessments of antioxidant potential in a wide variety of contexts and highlight the powerful protective properties of human seminal plasma.
2. Materials and Methods
2.1. Reagents and Suppliers
Unless otherwise stated, all reagents were purchased from Sigma Aldrich (St. Louis, MO, USA).
2.2. Human Sample Preparation
All human experiments were conducted following the protocols approved by the University of Newcastle Human Research and Ethics Committee and the State Government (H-2013-0319, H-2023-0385 and 200621). Semen samples obtained from healthy unselected male donors following 2-3 days of sexual abstinence and delivered to the laboratory within 1 hour of ejaculation. Whenever seminal plasma (SP) was required, the semen was centrifuged for 5 min at 500 × g and the plasma removed, either to be analyzed fresh, or after freezing in liquid nitrogen and storage at -80°C. Whenever spermatozoa were required, the semen was processed through Percoll gradients as previously described [18] and the high-density fraction resuspended in HEPES-buffered Biggers-Whitten-Whittingham medium (BWW) supplemented with 1 mg/mL polyvinyl alcohol at a concentration of 10 × 106/mL. In addition to spermatozoa, saliva samples were collected from randomly selected individuals and analyzed fresh without freezing.
2.3. Equine Sample Preparation
Institutional and New South Wales State Government ethical approval was secured for the use of equine material in this study (ACEC number A-2011-122). Equine semen was collected from normozoospermic Shetland and miniature crossbred pony stallions of proven fertility using a Missouri artificial vagina (AV; Minitube, Ballarat, VIC, Australia). Semen was centrifuged (5 min at 500 × g), and SP was collected. Blood was obtained with vacutainers using either a clotting tube for serum formation or a green-topped heparin tube for blood plasma. Tubes were centrifuged (10 min at 1000 × g) either immediately, or 10-15 min after collection in the case of serum, and the resulting supernatants frozen and stored at -80°C.
2.4. Bovine Sample Preparation
Institutional and New South Wales State Government ethical approval was secured for the use of bovine material in this study (ACEC number A-2022-223). The bulls utilized in this study were confined to a crush for semen collection. After a rectal examination to clear the anus of any excess fecal matter, a water-based, non-spermicidal lubricant was applied to a rectal probe (75 mm to 90 mm) and inserted into the anal cavity. Pulses from an electro-ejaculation unit were rhythmically applied (every 2-3 seconds, with 1-second interval breaks) until ejaculation occurred. A silicon funnel with a 15 mL Falcon tube attached was used to collect the ejaculate. The raw ejaculate was centrifuged (400 × g) for 20 min, and the SP collected, snap frozen and stored at -80°C. Cow ovaries were obtained from a local abattoir and transported back to the laboratory in prewarmed 0.9% saline solution (35-38°C). Antral follicles were subsequently aspirated using an 18-gauge needle attached to a 10 mL syringe and the resulting follicular fluid snap frozen in liquid nitrogen and stored at -80°C. In similar fashion, an 18-gauge needle was inserted into the bladder to allow the collection of post-mortem urine specimens, which were subsequently snap frozen and stored at -80°C.
2.5. Canine Sample Preparation
Canine semen was kindly collected and donated by an accredited breeder registered with Dogs NSW (Kerensa Kennels, Dogs NSW registration number 2100076617). Three healthy male dogs, 4–9 years old, were used for the semen collections. The breeds were an Australian Kelpie and two Border Collie dogs. The included ejaculates had > 80% morphologically normal spermatozoa and a motility of >80%. From each dog, the sperm rich fraction of the ejaculate was collected in a calibrated, plastic Falcon tube by digital manipulation [19]. A portion of each ejaculate was subsequently transferred to Eppendorf tubes and centrifuged at 500 × g for 5 min. The SP was then carefully collected from above the sperm pellet and transferred into clean tubes in 500 µL aliquots before being frozen and stored at -80°C.
2.6. Impact of cumene hydroperoxide on sperm motility
In order to determine whether the observed inter-species differences in levels of antioxidant protection provided by SP reflected the vulnerability of the spermatozoa to oxidative stress, time- and dose- dependent studies were conducted on the impact of cumene hydroperoxide on sperm motility. For this study spermatozoa from different species were prepared on discontinuous density gradients and the isolated spermatozoa suspended in medium at a concentration of 10 × 106/mL in BWW. Spermatozoa were then treated with various doses of cumene hydroperoxide (1, 0.5, 0.25, 0.125, 0.06 and 0 mM) and after two time points (15-30 min and 120-135 min) both total motility(average path velocity of > 5 µm/sec) and progressive motility (straightness of > 80% and average path velocity of > 25 µm/sec) were assessed using a CASA (Computer Aided Sperm Analysis) system (Hamilton Thorne, IVOS II, Beverly, MA, USA) and parameter settings that have been optimized for each species assessed.
In order to confirm the protective properties of human seminal plasma, Percoll-purified suspensions of human spermatozoa, at a concentration of 10 × 106/mL, were exposed cumene hydroperoxide (0.25 mM) in the presence or absence of varying amounts of human seminal plasma (12.5%, 6.25%, 3.125% and 0%). The impact of this treatment on motility was then determined using the above CASA system after 15-30 min exposure.
2.7. The antioxidant assay system
The RoXsta™ (Rapid Oxidative Stress Assays) antioxidant system used in these studies has been previously described in detail [17]. In essence, this assay uses ABTS as a redox active probe and measures 4 different kinds of antioxidant activity. (1) Organic peroxide scavenging activity, assessed by determining the ability of a given fluid to suppress the formation of the ABTS•+ radical using a reaction mixture containing 675 µL phosphate buffer, 25 µL ABTS (250 µM), 100 µL diluted sample and 100 µL hematin (0.05 mg/mL) at pH 6.5. The reaction was activated by the addition of 100 µL, 1 mM cumene hydroperoxide (100 µM final concentration) and incubated at room temperature for 20 min before the absorbance was determined at 734 nm in a spectrophotometer (SPECTROstar Nano, BMG Labtech). (2) Hydrogen peroxide scavenging activity, assessed by determining the ability of a given fluid to suppress formation of the ABTS•+ radical in a reaction mixture comprising 735 µL buffer, 15 µL ABTS (150 µM), 100 µL diluted sample and 50 µL HRP (0.05 mg/mL). In this case, the reaction was activated by the addition of 100 µL hydrogen peroxide (30 µM final concentration) and incubated at room temperature for 10 min to allow formation of the colored ABTS•+ radical cation which was then read at 734 nm. (3) Free radical scavenging activity was assessed by determining the ability of a given fluid to scavenge the ABTS•+ cation radical prepared by the oxidation of ABTS (100 µM) in phosphate buffer (pH 4.8) within the anodic chamber of an electrochemical cell [17]. For this post-activation assay, 1 mL of activated ABTS•+ was removed to a 1.5 mL Eppendorf tube and the suppression of absorbance measured at 734 nm after 5 min after addition of test sample in 15 µL. (4) Inhibition of free radical formation in a pre-activation assay in which 100 µL of diluted sample was added to 5.4 mL 100 µM ABTS at pH 4.8. A 2 mL aliquot of the diluted sample was then added to the anodic chamber of the electrochemical cell. Current (20 sec at 45V and 10 mA) was subsequently applied resulting in the appearance of ABTS•+ at the anode, the cathodic chamber serving as a passive control for sample turbidity. Following the passage of current, the anodic and cathodic chambers were agitated and then left for 5 min to allow the activation to proceed to completion and the suppression of ABTS•+ formation to be determined. For all of these antioxidant assays vitamin C (ascorbic acid) was used as a positive control and the results are expressed as vitamin C equivalents. Three repeat analyses were conducted on at least 3 independent biological replicates for each of the materials assessed in this study.
For each assay, optimization studies were conducted to determine the sample dilution at which the various assessments of antioxidant activity should be conducted. The results of this analysis are presented in Supplementary Table S1.
2.9. Statistics
Statical analysis of the data was undertaken using the JMP Pro 17 statistical software package (SAS Institute, Cary, NC, USA). All data are presented as the mean ± the standard error (S.E.M.) of at least 3 independent replicates. The data were initially screened using the Shapiro–Wilk test to determine the normality of data distribution. Data that did not conform to a normal distribution were transformed using the Box Cox transformation with optimized lambda values calculated for each parameter. In the Figures, all transformed data are presented in their original form for easier interpretation. Differences between groups were determined by ANOVA with Tukey HSD post hoc analysis. All significant differences were confirmed using non-parametric methods (Wilcoxon or Kruskal -Wallace tests). Throughout, the minimum level of statistical significance was set at p < 0.05.
3. Results
3.1. Impact of spermatozoa and freezing on the antioxidant activity of human semen
This analysis was initiated by an assessment of human semen samples to determine the levels of antioxidant activity expressed, and the extent to which they would be influenced by the presence or absence of spermatozoa and/or being frozen in liquid nitrogen (Figure 1A-D). The results indicated that for three of the antioxidant activities assessed (hydrogen peroxide scavenging, free radical scavenging and the suppression of free radical formation), the performance of human semen was neither impacted by the presence of spermatozoa nor being snap frozen in liquid nitrogen (Figure 1B-D). However, the organic peroxide scavenging assay revealed a modest decrease (p < 0.01) when semen was frozen and a slight reduction when spermatozoa were removed, that bordered statistical significance (p = 0.055) (Figure 1A).
Together, these results suggested that live spermatozoa make a minor contribution to the antioxidant capacity of human semen; a vast majority of the antioxidant activity was contained within the SP fraction and did not change significantly following freezing (Figure 1A). The dominant form of antioxidant activity in semen involved the capacity of this fluid to suppress free radical formation (~30-35 mM vitamin C equivalents), followed by organic peroxide scavenging (~13-15 mM vitamin C equivalents), hydrogen peroxide scavenging (~12-14 mM vitamin C equivalents) and free radical scavenging (0.75-1.35 mM vitamin C equivalents) activities (Figure 1A-D).
3.2. Antioxidant activity of semen relative to other biological fluids
In order to determine whether seminal plasma has evolved to provide spermatozoa with particularly high levels of antioxidant protection in relation to other biological fluids, the RoXsta™ system was used to undertake a comparative analysis of the antioxidant profiles expressed by blood, urine, saliva, ovarian follicular fluid and semen.
The results of this analysis revealed interesting differences between fluids in the levels of antioxidant protection afforded, as well as the types of antioxidant activity expressed (Figure 2A-D). With all four measures of antioxidant activity, human semen was significantly more active than any other fluid examined (p < 0.001) apart from urine, which was more active in scavenging free radicals and in suppressing the latter’s formation (Figure 2 C, D). By contrast, follicular fluid possessed significantly lower levels of antioxidant activity than semen suggesting that the ovarian follicle does not confer the same protection to oocytes as seminal plasma affords to spermatozoa (Figure 2A-D). Both blood serum and plasma exhibited low levels of antioxidant protection relative to urine and semen in all assays (p < 0001). Importantly however, some differences were observed between blood serum and plasma, that would influence the way in which the RoXsta™ system is applied in a clinical setting, since the optimum sampling strategy depended on the nature of the antioxidant activity being assessed. Thus, for the inhibition of free radical formation and organic peroxide scavenging activities, blood serum possessed significantly less activity than plasma (Figure 2A, C; p < 0.05), while for hydrogen peroxide scavenging activity, the opposite was the case (Figure 2B; p < 0.05). However, no significant differences were observed between serum and plasma when assessing free radical scavenging activity.
3.2. Seminal plasma in different species
Following on from these experiments, a comparative analysis was undertaken profiling the antioxidant activity of SP from different species (human, equine, bovine and canine SP). The results of this analysis indicated that the ability of human plasma to scavenge organic peroxides was significantly greater than that of any of the other species, while equine and bovine SPs possessed significantly (p < 0.001) more activity than their canine counterpart (Figure 3A). Similarly, in terms of hydrogen peroxide scavenging activity, human SP possessed significantly (p < 0.001) greater activity than any of the other species examined, while equine SP was significantly greater than bovine, and both equine and bovine SPs were more active than the canine material (p < 0.001) (Figure 3B).
The ability of human SP to suppress free radical formation was also significantly greater than either bovine or equine material (Figure 3 C; p < 0.001). However, with this assay, canine SP was just as active in the suppression of free radical formation as human SP and significantly greater than that observed with bovine or equine SP (Figure 3C; p < 0.001). Finally, the ability of human SP to scavenge free radicals was also higher (p < 0.001) than all of the other species examined, while both equine and bovine SP possessed more such activity than the canine equivalent (Figure 3D; p < 0.001). In general, the hierarchy of antioxidant protection observed in this comparative study was human > equine> bovine > canine, although the latter was surprisingly active in the suppression of free radical formation.
3.3. Impact of oxidative stress on sperm motility in different species
A possible explanation for the relatively high level of antioxidant protection afforded by human SP, particularly with respect to organic peroxide scavenging (Figure 2A) would be that human spermatozoa are particularly vulnerable to oxidative stress created by lipid peroxides. In order to test this hypothesis spermatozoa from three different species were exposed to cumene hydroperoxide and the impact on sperm motility assessed using a CASA system.
As hypothesised, human spermatozoa were found to be very vulnerable to oxidative attack. Thus, an analysis of sperm movement within 15-30 min of adding peroxide, revealed that both total and progressive motility were significantly (p < 0.001) reduced relative to control levels at doses of cumene hydroperoxide above 0.25 mM (Figure 3A). In contrast, no significant impact on bovine or equine spermatozoa was recorded under identical conditions (Figure 3C, E). When the exposure time to cumene hydroperoxide was extended to 2 h, the total and progressive motility of human spermatozoa was significantly suppressed at doses above 0.06 mM (p < 0.001), while at doses greater than 0.25 mM, these cells were completely motionless (Figure 3B). In the case of bovine spermatozoa however, neither the total nor the progressive motility of these cells was impacted by a 2h exposure to cumene hydroperoxide except at the highest dose examined of 1 mM (p < 0.05; Figure 3 D). Similarly, a 2 h exposure to cumene hydroperoxide did not impact the total motility of equine spermatozoa until doses of 0.5 mM (p < 0.01) and 1 mM (p < 0.001) were reached (Figure 3F), while progressive motility was not compromised until the cumene hydroperoxide dose exceeded 0.25 mM (p < 0.05; Figure 3 F). Taken together these data indicate a hierarchy of vulnerability such that human > equine > bovine, in keeping with the above analyses of SP antioxidant activity.
3.4. Impact of seminal plasma on peroxide-mediated toxicity
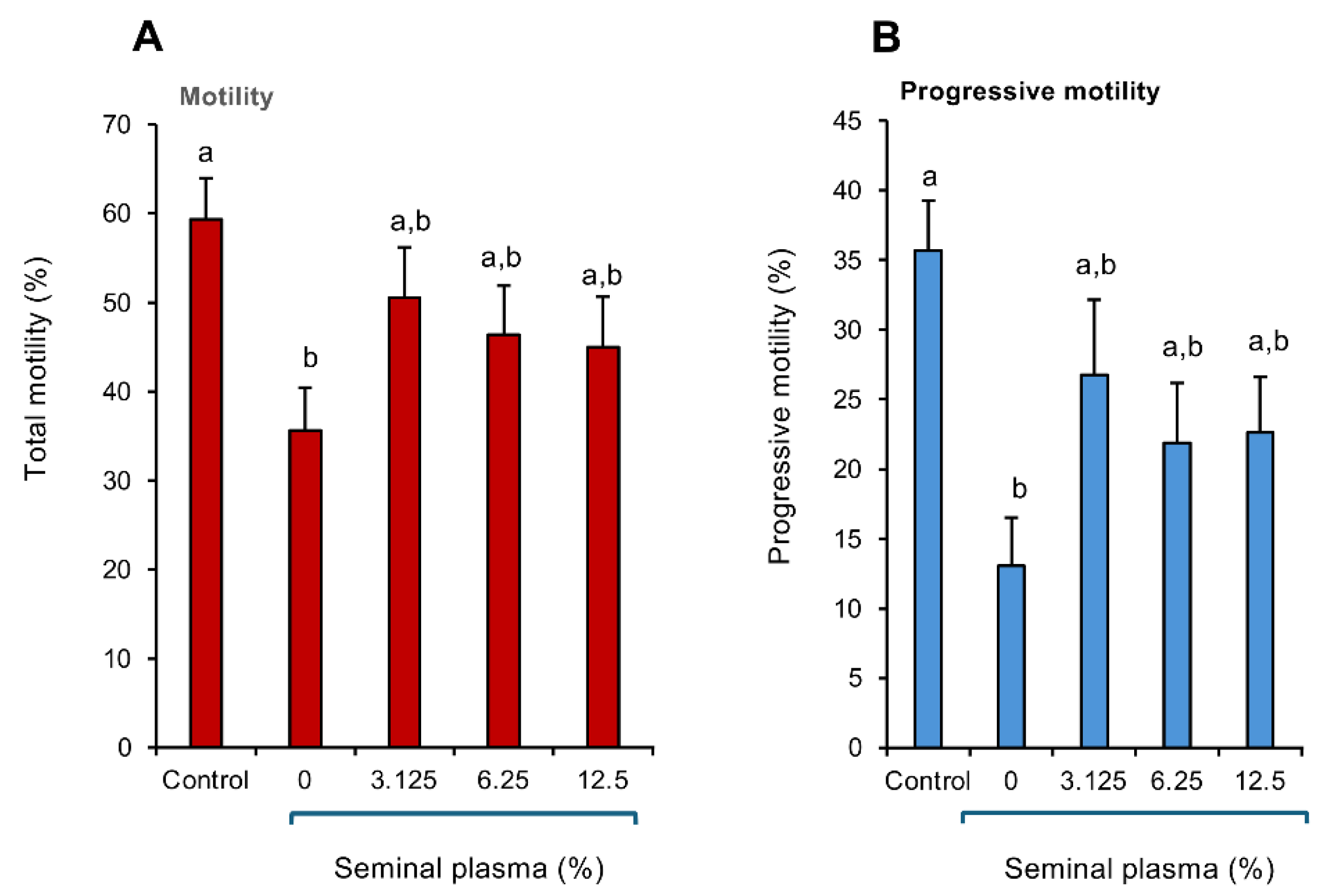
In order to confirm the powerful antioxidant protection offered by human seminal plasma, a study was performed in which the spermatozoa were exposed to cumene hydroperoxide in the presence or absence of human seminal plasma. Their functionality was then assessed with a focus on CASA measurements of total and progressive motility. This analysis revealed that exposure to 0.25 mM cumene peroxide precipitated the anticipated decline in both total (Figure 5A; p < 0.05) and progressive (Figure 5B; p < 0.01) motility within a 15-30 min time span (the time needed to conduct all of the CASA analyses). However, in the presence of as little as 3.125% human seminal plasma, this toxic oxidative impact was negated and neither the percentage motile nor progressively motile cells were different from the untreated control sample, as long as seminal plasma was present (Figure 5 A,B).
4. Discussion
The results of this study demonstrate that the RoXsta™ system is capable of rapidly and efficiently profiling antioxidant activity in a wide range of different biological materials, including blood serum and plasma, follicular fluid, saliva, urine and semen. One of the key features of this system is that it can be configured to measure different types of antioxidant activity, not just the ability of a given fluid to act as a free radical scavenger. In the conventional ABTS-mediated Total Antioxidant Capacity (TAC) assay, fluids are assessed for their ability to neutralise the preformed ABTS•+ radical cation, which is generated using powerful oxidizing agents such as potassium persulphate or the combination of metmyoglobin and hydrogen peroxide [17]. With the Roxsta™ system, the ABTS•+ radical is generated electrochemically, permitting determination of the ability of a sample to either suppress ABTS•+ free radical formation or scavenge this radical, depending on whether the test material is added before or after the application of current. Alternatively, the oxidation of ABTS can be catalysed using combinations of hydrogen peroxide and HRP or cumene hydroperoxide and hematin, thereby permitting the rapid assessment of hydrogen peroxide- and organic peroxide- scavenging activities respectively [17].
This study has clearly demonstrated the versatility of the RoXsta™ system in characterizing the antioxidant profile of complex biological mixtures. To illustrate this point, we have shown that this system is not only competent to assess the relative antioxidant protection offered by mammalian body fluids, but is also capable of profiling plant materials demonstrating, for example, the relative superiority of pomegranate juice compared with orange, grape, super berry, cranberry, and cherry juices (Figure S1, supplementary materials). We have further demonstrated that this system is effective in differentiating the relative antioxidant activity of non-biological products such as commercial face sera, revealing the existence of significant variation between commercial cosmetic products in the type of antioxidant protection offered (Figure S2, supplementary materials). Thus, this system can profile the antioxidant activity exhibited a wide range of different starting materials, much faster than any other methodology described to date.
In the comparison of different biofluids we were naturally limited to the biofluids that were available to us including equine blood plasma and serum, human saliva and semen and bovine follicular fluid and urine. Despite the use of different source species, the results are generally representative of the fluid in question. Thus, our free radical scavenging assay is broadly similar to the post-activation TAC assay originally developed by Rice-Evans and colleagues except that the ABTS•+ radical is generated electrochemically rather than using the conventional potassium persulphate or metmyoglobin/H2O2 ABTS oxidation strategy [20]. Despite these methodological differences, our assessment of the antioxidant potential of human seminal plasma (Figure 2D) or intact semen (Figure 2D) using the free radical scavenging assay (~1.4 mM vitamin C equivalents) agrees extremely well with the value of 1.4 mM Trolox equivalents generated by Mahfouz et al. [21] using the conventional TAC assay. Similarly, our estimation of salivary antioxidant levels using the free radical scavenging assay (~0.22 mM vitamin C equivalents) approximates to published values for human saliva acquired using the traditional ABTS assay (0.34 ± 0.037 mM Trolox equivalents) [22] and agrees well with similar estimates made in pigs (~0.3-0.4 mM Trolox equivalents) [23]. In terms of blood, our estimates of the free radical scavenging activity of equine plasma and serum (~0.41 and 0.39 mM vitamin C equivalents respectively) agrees with previous assessments of equine blood plasma (~0.37 - 0.40 mM Trolox equivalents) using the ABTS-based TAC assay [24] and reflects similar determinations in the dog (0.312 ± 0.015 mM Trolox equivalents) [25], cattle (~0.12 mM Trolox equivalents) [26] and humans (0.29 mM Trolox equivalents) [27]. Similarly, the intense antioxidant activity measured in bovine urine with the free radical scavenging assay (~4 mM Vitamin C equivalents) reflects the high values recorded in human (3.13 ± 0.11 mM Trolox equivalents) with ABTS free radical scavenging assays [28] and comparative studies conducted using alternative methods to measure total antioxidant activity in human body fluids [29]. Our finding that follicular fluid possesses a relatively low free radical scavenging capacity (generally < 1.5 mM) compared with semen and urine, resonates with previous analyses on water buffalo [30], humans [31] and cattle [32].
The comparative analysis of biofluids also emphasized the high antioxidant properties of urine, presumably as a result of the latter’s uric acid content, confirming the results of other comparative studies [29]. The intrinsic antioxidant activity of urine appeared to be particularly evident in the context of free radical scavenging and inhibition of ABTS•+ radical formation, which involves the mediation of hydroxyl radicals and is in keeping with the known capacity of uric acid to scavenge this radical species [33,34]. In contrast, urine was relatively less active in the hydrogen peroxide scavenging assay, possibly because uric acid reacts poorly with this oxidant [35].
Naturally, blood plasma or serum are recognized as matrices for the determination of total antioxidant activity. With the RoXsta™ system both plasma and serum appeared to be equally effective in reflecting the organic peroxide-, hydrogen peroxide- and free radical- scavenging activity of blood. The exception was the inhibition of free radical formation assay which not only detected much higher concentrations of antioxidant activity in blood than the other assays but also revealed a significant difference between serum and plasma with the latter presenting significantly higher levels of activity than the former (Figure 2C). Evidently the blood-borne factors responsible for suppressing formation of the ABTS•+ radical in an electrochemical cell, can be compromised by the clotting procedures used to generate serum. This could be due to the degradation of existing antioxidants in blood, such as vitamin C, or possibly the enhanced consumption of antioxidants due to the activation of ROS generation by platelets during the clotting process [36].
Notwithstanding the significant antioxidant activity present in blood and urine, the most powerful antioxidant activity was consistently observed in human semen. In order to determine whether this was a unique feature of human semen or a general characteristic of SP, we examined semen samples from 4 different sources (human, bovine, equine and canine) and revealed hitherto unknown, and unsuspected, interspecies differences in the level and type of antioxidant protection afforded by this fluid. In general, human seminal plasma was shown to possess the greatest antioxidant potential, particularly in terms of its organic-, and hydrogen-, peroxide scavenging activity. This observation accords with existing data indicating that human spermatozoa are particularly susceptible to the toxic impact of lipid peroxides and that this impact can be reversed by the addition of SP [37]. These data also accord with a large number of studies indicating that antioxidant capacity of human seminal plasma is significantly lower in cases of male infertility and is inversely correlated with the accumulation of lipid peroxidation markers such as malondialdehyde [38,39]. Analyses of the TAC values associated with human semen have also shown positive correlations with multiple parameters of semen quality including, sperm motility, concentration, morphology and DNA integrity [40,41,42], while being negatively associated with measurements of lipid peroxidation [38]. Furthermore, the losses of sperm function associated with clinical conditions such as varicocele [43], environmental exposures to, for example, air pollutants and glyphosate [44], and lifestyle factors, including cigarette smoking, diabetes, and obesity [45,46], are all associated with decreases in the total antioxidant protection offered by SP.
Most of these studies have been conducted using the ABTS-based TAC assay and therefore reflect the ability of SP to scavenge the ABTS•+ radical. However, others have also measured hydrogen peroxide scavenging activity in assays exploiting the HRP-mediated oxidation of luminol [47]. Recently, electron spin resonance spectroscopy was used to monitor the radical scavenging activity of seminal plasma from multiple species (stallion, bull, man and lion) [48]. This analysis also revealed that the free radical scavenging capacity human SP was more than ten times more effective than the other species examined, in accord with the data presented in the present analysis [48]. Furthermore, it indicated the presence of significant free radical scavenging activity in bovine SP, in keeping with the present study and the result obtained by others, who detected antioxidant activity in bovine ejaculates that exhibited the anticipated negative correlation with lipid peroxidation status [49]. However, the study by Jakop et al. [48] failed to detect significant antioxidant activity in equine SP, in contrast to our data, and the results of others who have found significant free radical scavenging activity in equine SP that had a significant impact on sperm function and DNA integrity [50]. For its part, canine SP has also been shown to possess a capacity for ABTS•+ radical scavenging by our group and others [51]. However, this is the first study to reveal the relative paucity of this activity in canine semen compared with other species in terms of its organic peroxide-, hydrogen peroxide- and free radical- scavenging activity. It is also the first to reveal that canine SP possesses a particularly high capacity to suppress free radical formation (Figure 3C).
The relatively high level of antioxidant protection afforded by human SP emphasises the importance that evolution has attached to protecting human spermatozoa from oxidative stress. In light of these findings, the RoXsta™ system may be of value in rapidly diagnosing antioxidant deficiencies within the patient population and identifying those individuals for whom antioxidant therapy represents a rational therapeutic option. [16]. Interestingly, the egg is not offered the same high level of antioxidant protection given the relatively modest levels of antioxidant activity detected in follicular fluid, which was not fundamentally higher than blood plasma, with the exception of the inhibition of free radical formation where it was lower (Figure 2C). Although relatively low, the levels of free radical scavenging activity measured in follicular fluid correspond with those observed in clinical studies (~0.3 - 0.7 mM) which have, in turn, been correlated with the incidence of pregnancy in assisted conception cycles [52].
The need for greater extracellular antioxidant protection for spermatozoa compared with oocytes is entirely explicable on the basis of cell architecture. Spermatozoa possess an extremely limited cytoplasmic space in terms of absolute volume and restricted location within the sperm midpiece, as a result of which there is little space in which to house the antioxidant factors that protect most cells from oxidative stress. Consequently, these cells are uniquely dependent on the availability of antioxidants in the extracellular space during their passage from the germinal epithelium of the testes to the ampulla of the fallopian tubes, where fertilization occurs. It is for this reason that SP, as well as the luminal spaces inhabited by spermatozoa within the testes, epididymis, and female tract are so well endowed with antioxidants [53,54,55]. By contrast, the oocyte possesses significant cytoplasmic volume in which to sequester cellular antioxidants and, in any case, is metabolically dormant for much of its life, surrounded by a protective cocoon of pre-granulosa cells within the primordial follicle [56]. However, as soon as the oocyte starts to grow and mature within the Graafian follicle, it becomes vulnerable to oxidative stress and dependent on the antioxidant protection afforded by follicular fluid [57,58], even if such protection does not approach the level of antioxidant protection offered to spermatozoa by SP.
The particularly high levels of antioxidant protection offered by human SP in comparison to other species reflects the particular vulnerability of human spermatozoa to peroxide attack as revealed in Figure 4 A-D and Figure 5 A,B. This vulnerability may stem from differences in the energy metabolism exhibited by the spermatozoa of different species. Thus canine, equine and bovine spermatozoa are heavy users of oxidative phosphorylation [59,60]. As a consequence, ROS generation is a normal feature of sperm metabolism in these species to the extent that positive correlations have been observed between sperm motility and intracellular ROS generation [60,61]. Given this inherently high level of exposure to ROS, mechanisms have evolved to provide some level of resistance against oxidative stress in those species that exploit the high levels of ATP generation associated with intense mitochondrial respiration [62]. By contrast, human spermatozoa are built more for distance than for speed. This strategy may have evolved because our species is unusual in that insemination is not synchronized with ovulation through the mediation of behavioural oestrus. As a result, human spermatozoa may have to survive in the female tract for several days before an egg is released from the ovary. To achieve this longevity, human spermatozoa have developed a dependency on glycolysis, sacrificing the ATP generating capacity of oxidative phosphorylation for a low level of exposure to mitochondrial ROS [63,64]. This lack of exposure to intracellular ROS has created a dependency on antioxidant protection in the extracellular space to defend these cells against oxidative stress; hence the unusually high levels of antioxidant protection offered by human SP, and its diagnostic significance.
5. Conclusions
In conclusion, the RoXsta™ system appears to be a convenient rapid method of measuring and characterising antioxidant activity in a wide variety of clinical and commercial contexts. The simplicity and rapidity of the RoXsta™ system should be of value to clinicians since it provides an opportunity for real time point-of-care assessments to diagnose those patients suffering from oxidative stress and to monitor the effectiveness of antioxidant supplementation in restoring an appropriate redox balance. Such assessments may be particularly important in the field of male infertility where deficiencies in the levels of antioxidant afforded to the spermatozoa have been clearly, and causally, linked to the impairment of human sperm function.
6. Patents
The methodology described in the article is subject to a patent application (PCT/AU2024/050943).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1 Optimised sample dilutions for antioxidant assays; Figure S1 Ability of the RoXsta™ system to profile the antioxidant activity of fruit juices. Figure S2: Ability of the RoXsta™ system to profile the antioxidant activity of cosmetic products.
Author Contributions
Conceptualization, R.J.A.; methodology, A.W., N.H., M.B., K.M., S.L.; formal analysis, R.J.A.; investigation, A.W., N.H., M.B., K.M., S.L.; resources, R.J.A.; data curation, A.W., N.H.; writing—original draft preparation, R.J.A.; writing—review and editing, A.W., N.H., M.B., S.L., Q.V.; project administration, R.J.A.; funding acquisition, R.J.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Memphasys Ltd in a grant to the University of Newcastle, grant number G2201336.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Boards for Human and Animal Ethics at the University of Newcastle. For studies involving human subjects the project was approved by both the University of Newcastle Human Research and Ethics Committee and the State Government (H-2013-0319 and 200621). For studies involving equine material, Institutional and New South Wales State Government ethical approval was secured, ACEC number A-2011-122, while for bovine material, the approval number was ACEC number A-22-223.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data are contained within the article.
Acknowledgments
We wish to thank the University of Newcastle for the facilities to conduct this study. We also thank the Arrowfield stud, Segenhoe, NSW for the supply of equine blood samples.
Conflicts of Interest
R.J.A. is a scientific advisor to Memphasys Ltd, which funded the project in a collaboration in the University of Newcastle. NH, AW, MB and SL are funded by a grant to the University of Newcastle from Memphasys Ltd (Grant number G2201336). The funders played no role in the detailed design of the study or in the collection, the analyses or interpretation of the data or in the writing of the manuscript. However, they were supportive of the desire to publish these results.
References
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A. The role of genetics and oxidative stress in the etiology of male infertility-a unifying hypothesis? Front. Endocrinol. 2020, 11, 581838. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H. . Targeting oxidative stress in disease: promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Steiner, A.Z.; Hansen, K.R.; Barnhart, K.T.; Cedars, M.I.; Legro, R.S.; Diamond, M.P.; Krawetz, S.A.; Usadi, R.; Baker, V.L.; Coward, R.M.; et al. The effect of antioxidants on male factor infertility: the Males; Antioxidants; and Infertility (MOXI) randomized clinical trial. Fertil. Steril. 2020, 113, 552–560.e3. [Google Scholar] [CrossRef]
- Ma, L.; Sun, Y. Comparison of L-Carnitine vs. Co Q10 and Vitamin E for idiopathic male infertility: a randomized controlled trial. Eur. Rev. Med Pharmacol. Sci. 2022, 26, 4698–4704. [Google Scholar]
- Zhang, Q.F.; Wang, S.; Zhang, H.; Liu, Q.L.; Wei, Y.; Deng, W.; Wang, C.; Yang, B. Effects of alpha-lipoic acid on sperm quality in patients with varicocele-related male infertility: study protocol for a randomized controlled clinical trial. Trials 2022, 23, 1002. [Google Scholar] [CrossRef]
- Jannatifar, R.; Parivar, K.; Roodbari, N.H.; Nasr-Esfahani, M.H. Effects of N-acetyl-cysteine supplementation on sperm quality; chromatin integrity and level of oxidative stress in infertile men. Reprod. Biol. Endocrinol. 2019, 17, 24. [Google Scholar] [CrossRef]
- Ghanem, H.; Shaeer, O.; El-Segini, A. Combination clomiphene citrate and antioxidant therapy for idiopathic male infertility: a randomized controlled trial. Fertil. Steril. 2010, 93, 2232–2235. [Google Scholar] [CrossRef]
- ElSheikh, M.G.; Hosny, M.B.; Elshenoufy, A.; Elghamrawi, H.; Fayad, A.; Abdelrahman, S. Combination of vitamin E and clomiphene citrate in treating patients with idiopathic oligoasthenozoospermia: A prospective; randomized trial. Andrology 2015, 3, 864–867. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Shafiei, N.; Safarinejad, S. A prospective double-blind randomized placebo-controlled study of the effect of saffron (Crocus sativus Linn.) on semen parameters and seminal plasma antioxidant capacity in infertile men with idiopathic oligoasthenoteratozoospermia. Phytother. Res. 2011, 25, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Comhaire, F.H.; El Garem, Y.; Mahmoud, A.; Eertmans, F.; Schoonjans, F. Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: a double blind; randomized trial. Asian J. Androl. 2005, 7, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Habibi, M.; Fakhari Zavareh, Z.; Abbasi, B.; Esmaeili, V.; Shahverdi, A.; Sadighi Gilani, M.A.; Tavalaee, M.; Nasr-Esfahani, M.H. Alpha-lipoic acid supplementation for male partner of couples with recurrent pregnancy loss: a post hoc analysis in clinical trial. Int. J. Fertil. Steril. 2023, 17, 67–74. [Google Scholar]
- de Ligny, W.; Smits, R.M.; Mackenzie-Proctor, R.; Jordan, V.; Fleischer, K.; de Bruin, J.P.; Showell, M.G. Antioxidants for male subfertility. Cochrane Database Syst. Rev. 2022, 5, CD007411. [Google Scholar]
- Sengupta, P.; Dutta, S.; Alahmar, A.T. reductive stress and male infertility. Adv. Exp. Med. Biol. 2022, 1391, 311–321. [Google Scholar] [PubMed]
- Suleiman, S.A.; Ali, M.E.; Zaki, Z.M.; el-Malik, E.M.; Nasr, M.A. Lipid peroxidation and human sperm motility: protective role of vitamin E. J. Androl. 1996, 17, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Antioxidant trials-the need to test for stress. Hum. Reprod. Open 2021, 2021, hoab007. [Google Scholar] [CrossRef]
- Aitken, R.J.; Wilkins, A.; Harrison, N.; Kobarfard, K.; Lambourne, S. Development of novel; rapid point-of-care assays for monitoring different forms of antioxidant activity – the RoXsta™ system. Antioxidants 2024.
- Aitken, R.J.; Buckingham, D.W.; Harkiss, D.; Paterson, M.; Fisher, H.; Irvine, D.S. The extragenomic action of progesterone on human spermatozoa is influenced by redox regulated changes in tyrosine phosphorylation during capacitation. Mol. Cell. Endocrinol. 1996, 117, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Linde-Forsberg, C. Artificial insemination with fresh; chilled extended and frozen-thawed semen in the dog. Sem. Vet. Med. Surg. 1995, 10, 48–58. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Mahfouz, R.; Sharma, R.; Sharma, D.; Sabanegh, E.; Agarwal, A. Diagnostic value of the total antioxidant capacity (TAC) in human seminal plasma. Fertil. Steril. 2009, 91, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Tarboush, N.A.; Al Masoodi, O.; Al Bdour, S.; Sawair, F.; Hassona, Y. Antioxidant capacity and biomarkers of oxidative stress in saliva of khat-chewing patients: a case-control study. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 127, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.P.; Mainau, E.; Cerón, J.J.; Contreras-Aguilar, M.D.; Martínez-Subiela, S.; Navarro, E.; Tecles, F.; Manteca, X.; Escribano, D. Biomarkers of oxidative stress in saliva in pigs: analytical validation and changes in lactation. BMC Vet. Res. 2019, 15, 144. [Google Scholar] [CrossRef] [PubMed]
- Ott, E.C.; Cavinder, C.A.; Wang, S.; Smith, T.; Lemley, C.O.; Dinh, T.T.N. Oxidative stress biomarkers and free amino acid concentrations in the blood plasma of moderately exercised horses indicate adaptive response to prolonged exercise training. J. Anim. Sci. 2022, 100, skac086. [Google Scholar] [CrossRef] [PubMed]
- Michałek, M.; Tabiś, A.; Noszczyk-Nowak, A. Serum total antioxidant capacity and enzymatic defence of dogs with chronic heart failure and atrial fibrillation: a preliminary study. J. Vet. Res. 2020, 64, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.K.; Singh, S.K.; Nakade, U.P.; Singh, V.K.; Sharma, A.; Srivastava, M.; Yadav, B.; Singh, Y.; Sirohi, R.; Garg, S.K. Ameliorative potential of prepartal trace mineral and vitamin supplementation on parturition-induced redox balance and myeloperoxidase activity of periparturient sahiwal cows. Biol. Trace Elem. Res. 2017, 177, 72–79. [Google Scholar] [CrossRef]
- Horváth-Szalai, Z.; Jakabfi-Csepregi, R.; Szirmay, B.; Ragán, D.; Simon, G.; Kovács-Ábrahám, Z.; Szabó, P.; Sipos, D.; Péterfalvi, Á.; Miseta, A.; Csontos, C.; et al. Serum total antioxidant capacity (TAC) and TAC/Lymphocyte ratio as promising predictive markers in COVID-19. Int. J. Mol. Sci. 2023, 24, 12935. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Chia, M. Urinary total antioxidant capacity in soccer players. Acta Kinesiologica. 2009, 3, 26–33. [Google Scholar]
- Ziobro, A.; Bartosz, G. A comparison of the total antioxidant capacity of some human body fluids. Cell. Mol. Biol Lett. 2003, 8, 415–419. [Google Scholar]
- Kosior, M.A.; Esposito, R.; Cocchia, N.; Piscopo, F.; Longobardi, V.; Cacciola, N.A.; Presicce, G.A.; Campanile, G.; Aardema, H.; Gasparrini, B. Seasonal variations in the metabolomic profile of the ovarian follicle components in Italian Mediterranean Buffaloes. Theriogenology 2023, 202, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Oyawoye, O.A.; Abdel-Gadir, A.; Garner, A.; Leonard, A.J.; Perrett, C.; Hardiman, P. The interaction between follicular fluid total antioxidant capacity; infertility and early reproductive outcomes during in vitro fertilization. Redox Rep. 2009, 14, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Choi, A.; Yu, H.Y.; Czerniak, S.M.; Holick, E.A.; Paolella, L.J.; Agarwal, A.; Combelles, C.M. Fluctuations in total antioxidant capacity; catalase activity and hydrogen peroxide levels of follicular fluid during bovine folliculogenesis. Reprod. Fertil. Dev. 2011, 23, 73–80. [Google Scholar] [CrossRef]
- Becker, B.F.; Reinholz, N.; Ozçelik, T.; Leipert, B.; Gerlach, E. Uric acid as radical scavenger and antioxidant in the heart. Pflugers Arch. 1989, 415, 127–135. [Google Scholar] [CrossRef]
- Cohen, A.M.; Aberdroth, R.E.; Hochstein, P. Inhibition of free radical-induced DNA damage by uric acid. FEBS Lett. 1984, 174, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Halliwell, B. Action of biologically-relevant oxidizing species upon uric acid. Identification of uric acid oxidation products. Chem. Biol. Interact. 1990, 73, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Ialongo, C. Preanalytic of total antioxidant capacity assays performed in serum; plasma; urine and saliva. Clin. Biochem. 2017, 50, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Mann, T.; Sherins, R. Peroxidative breakdown of phospholipids in human spermatozoa; spermicidal properties of fatty acid peroxides; and protective action of seminal plasma. Fertil. Steril. 1979, 31, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, S.; Tagliamonte, M.C.; Catalani, S.; Primiterra, M.; Canestrari, F.; De Stefani, S.; Palini, S.; Bulletti, C. Differences in blood and semen oxidative status in fertile and infertile men; and their relationship with sperm quality. Reprod. Biomed. Online 2012, 25, 300–306. [Google Scholar] [CrossRef]
- Smith, R.; Vantman, D.; Ponce, J.; Escobar, J.; Lissi, E. Total antioxidant capacity of human seminal plasma. Hum. Reprod. 1996, 11, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Khosrowbeygi, A.; Zarghami, N. Levels of oxidative stress biomarkers in seminal plasma and their relationship with seminal parameters. BMC Clin. Pathol. 2007, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Pahune, P.P.; Choudhari, A.R.; Muley, P.A. The total antioxidant power of semen and its correlation with the fertility potential of human male subjects. J. Clin. Diagn Res. 2013, 7, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, F.; Valojerdi, M.R.; Amanlou, M.; Karimian, L.; Abolhassani, F. Relationship of seminal reactive nitrogen and oxygen species and total antioxidant capacity with sperm DNA fragmentation in infertile couples with normal and abnormal sperm parameters. Andrologia 2014, 46, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Giulini, S.; Sblendorio, V.; Xella, S.; La Marca, A.; Palmieri, B.; Volpe, A. Seminal plasma total antioxidant capacity and semen parameters in patients with varicocele. Reprod. Biomed. Online 2009, 18, 617–621. [Google Scholar] [CrossRef]
- Vasseur, C.; Serra, L.; El Balkhi, S.; Lefort, G.; Ramé, C.; Froment, P.; Dupont, J. Glyphosate presence in human sperm: First report and positive correlation with oxidative stress in an infertile French population. Ecotoxicol. Environ. Saf. 2024, 278, 116410. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.A.; Agarwal, A.; Sharma, R.K.; Nelson, D.R.; Thomas, A.J. Jr. Effect of cigarette smoking on levels of seminal oxidative stress in infertile men: a prospective study. Fertil. Steril. 2002, 78, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Abbasihormozi, S.H.; Babapour, V.; Kouhkan, A.; Niasari Naslji, A.; Afraz, K.; Zolfaghary, Z.; Shahverdi, A.H. Stress hormone and oxidative stress biomarkers link obesity and diabetes with reduced fertility potential. Cell J. 2019, 21, 307–313. [Google Scholar] [PubMed]
- Said, T.M.; Kattal, N.; Sharma, R.K.; Sikka, S.C.; Thomas, A.J., Jr.; Mascha, E.; Agarwal, A. Enhanced chemiluminescence assay vs colorimetric assay for measurement of the total antioxidant capacity of human seminal plasma. J. Androl. 2003, 24, 676–680. [Google Scholar] [CrossRef]
- Jakop, U.; Müller, K.; Müller, P.; Neuhauser, S.; Callealta Rodríguez, I.; Grunewald, S.; Schiller, J.; Engel, K.M. Seminal lipid profiling and antioxidant capacity: A species comparison. PLoS One 2022, 17, e0264675. [Google Scholar] [CrossRef]
- Gürler, H.; Calisici, O.; Bollwein, H. Inter- and intra-individual variability of total antioxidant capacity of bovine seminal plasma and relationships with sperm quality before and after cryopreservation. Anim. Reprod. Sci. 2015, 155, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.A.; Matás, C.; Torrecillas, A.; Saez, F.; Gadea, J. Seminal plasma components from fertile stallions involved in the epididymal sperm freezability. Andrology 2021, 9, 728–743. [Google Scholar] [CrossRef] [PubMed]
- Risso, A.; Pellegrino, F.J.; Corrada, Y.; Schinella, G. Evaluation of total antioxidant activity and oxidative stress in seminal plasma from dogs supplemented with fish oil and vitamin E. Int. J. Fertil. Steril. 2021, 15, 15–19. [Google Scholar]
- Afrough, M.; Nikbakht, R.; Hashemitabar, M.; Ghalambaz, E.; Amirzadeh, S.; Zardkaf, A.; Adham, S.; Mehdipour, M.; Dorfeshan, P. Association of follicular fluid antioxidants activity with aging and in vitro fertilization outcome: a cross-sectional study. Int. J. Fertil. Steril. 2024, 18, 115–122. [Google Scholar] [PubMed]
- Aitken, R.J.; Roman, S.D. Antioxidant systems and oxidative stress in the testes. Oxid. Med. Cell Longev. 2008, 1, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Vernet, P.; Aitken, R.J.; Drevet, J.R. Antioxidant strategies in the epididymis. Mol. Cell. Endocrinol. 2004, 216, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Rahiminejad, M.E.; Moaddab, A.; Ganji, M.; Eskandari, N.; Yepez, M.; Rabiee, S.; Wise, M.; Ruano, R.; Ranjbar, A. Oxidative stress biomarkers in endometrial secretions: A comparison between successful and unsuccessful in vitro fertilization cycles. J. Reprod. Immunol. 2016, 116, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Ernst, E.H.; Lykke-Hartmann, K. Transcripts encoding free radical scavengers in human granulosa cells from primordial and primary ovarian follicles. J. Assist. Reprod. Genet. 2018, 35, 1787–1798. [Google Scholar] [CrossRef]
- Pasqualotto, E.B.; Agarwal, A.; Sharma, R.K.; Izzo, V.M.; Pinotti, J.A.; Joshi, N.J.; Rose, B.I. Effect of oxidative stress in follicular fluid on the outcome of assisted reproductive procedures. Fertil. Steril. 2004, 81, 973–976. [Google Scholar] [CrossRef]
- Martin, J.H.; Nixon, B.; Café, S.L.; Aitken, R.J.; Bromfield, E.G.; Lord, T. Oxidative stress and in vitro ageing of the post-ovulatory oocyte: an update on recent advances in the field. Reproduction 2022, 164, F109–F124. [Google Scholar] [CrossRef]
- Peña, F.J.; Ortiz-Rodríguez, J.M.; Gaitskell-Phillips, G.L.; Gil, M.C.; Ortega-Ferrusola, C.; Martín-Cano, F.E. An integrated overview on the regulation of sperm metabolism (glycolysis-Krebs cycle-oxidative phosphorylation). Anim. Reprod. Sci. 2022, 246, 106805. [Google Scholar] [CrossRef]
- Foutouhi, A.; Meyers, S. Comparative oxidative metabolism in mammalian sperm. Anim. Reprod Sci. 2022, 247, 107095. [Google Scholar] [CrossRef]
- Gibb, Z.; Lambourne, S.R.; Aitken, R.J. The paradoxical relationship between stallion fertility and oxidative stress. Biol. Reprod. 2014, 91, 77. [Google Scholar] [CrossRef]
- Gibb, Z.; Lambourne, S.R.; Curry, B.J.; Hall, S.E.; Aitken, R.J. Aldehyde dehydrogenase plays a pivotal role in the maintenance of stallion sperm motility. Biol. Reprod. 2016, 94, 133. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.C.; Ford, W.C. The role of glucose in supporting motility and capacitation in human spermatozoa. J. Androl. 2001, 22, 680–695. [Google Scholar] [CrossRef]
- Hereng, T.H.; Elgstøen, K.B.; Cederkvist, F.H.; Eide, L.; Jahnsen, T.; Skålhegg, B.S.; Rosendal, K.R. Exogenous pyruvate accelerates glycolysis and promotes capacitation in human spermatozoa. Hum. Reprod. 2011, 26, 3249–3263. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Impact of sperm presence and freezing on the antioxidant activity of human semen. (A) Organic peroxide scavenging activity. (B) Hydrogen peroxide scavenging activity. (C) Inhibition of ABTS•+ radical formation. (D) ABTS•+ radical scavenging activity. All results are expressed as vitamin C equivalents. Statistical analysis by ANOVA and confirmed with non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; (n = 9).
Figure 1.
Impact of sperm presence and freezing on the antioxidant activity of human semen. (A) Organic peroxide scavenging activity. (B) Hydrogen peroxide scavenging activity. (C) Inhibition of ABTS•+ radical formation. (D) ABTS•+ radical scavenging activity. All results are expressed as vitamin C equivalents. Statistical analysis by ANOVA and confirmed with non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; (n = 9).
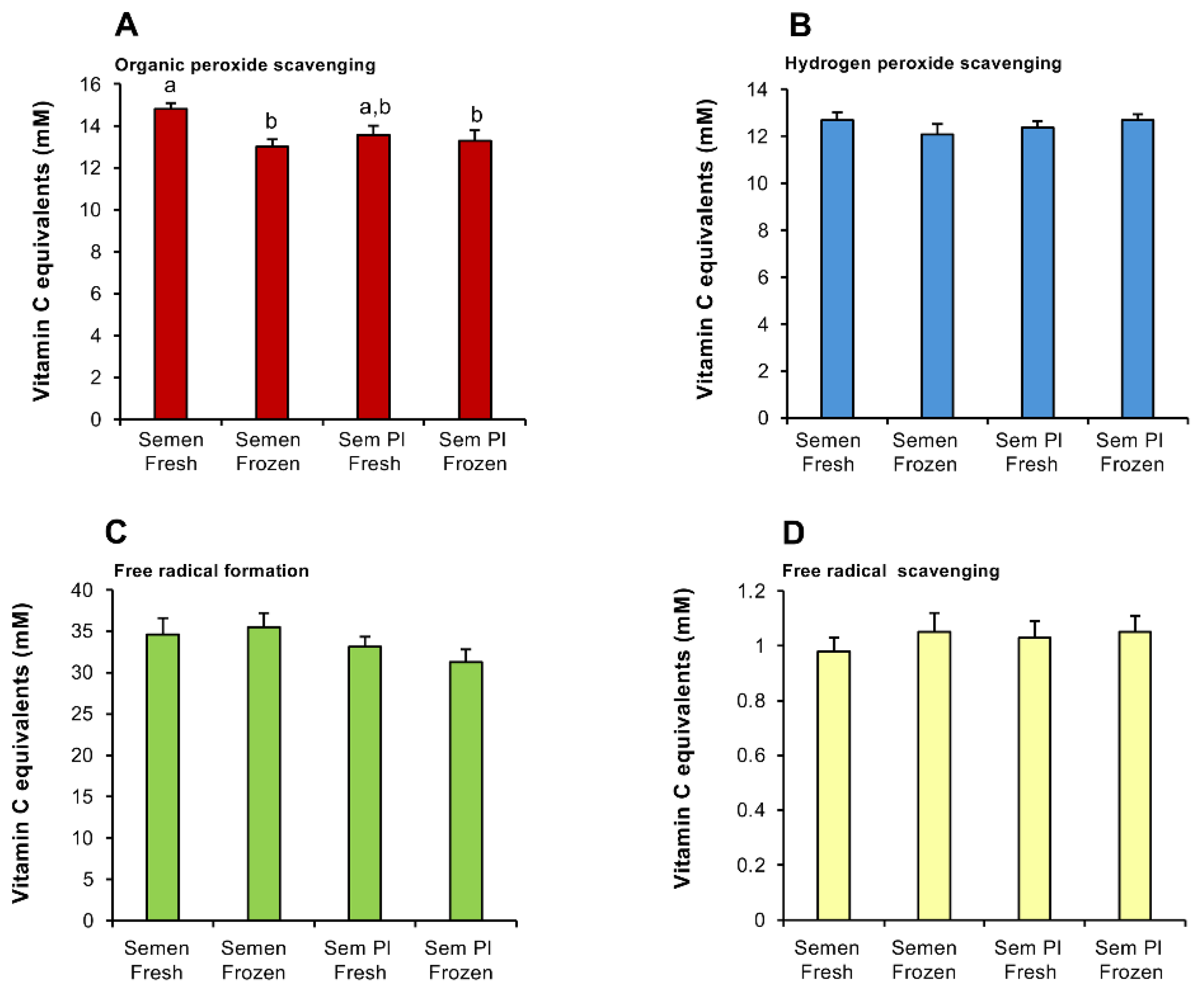
Figure 2.
Analysis of antioxidant activity in a range of biofluids using the RoXsta™ system. (A) Organic peroxide scavenging activity. (B) Hydrogen peroxide scavenging activity. (C) Inhibition of ABTS•+ radical formation. (D) ABTS•+ radical scavenging activity. All results are expressed as vitamin C equivalents. Statistical analysis by ANOVA and confirmed using non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 9.
Figure 2.
Analysis of antioxidant activity in a range of biofluids using the RoXsta™ system. (A) Organic peroxide scavenging activity. (B) Hydrogen peroxide scavenging activity. (C) Inhibition of ABTS•+ radical formation. (D) ABTS•+ radical scavenging activity. All results are expressed as vitamin C equivalents. Statistical analysis by ANOVA and confirmed using non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 9.
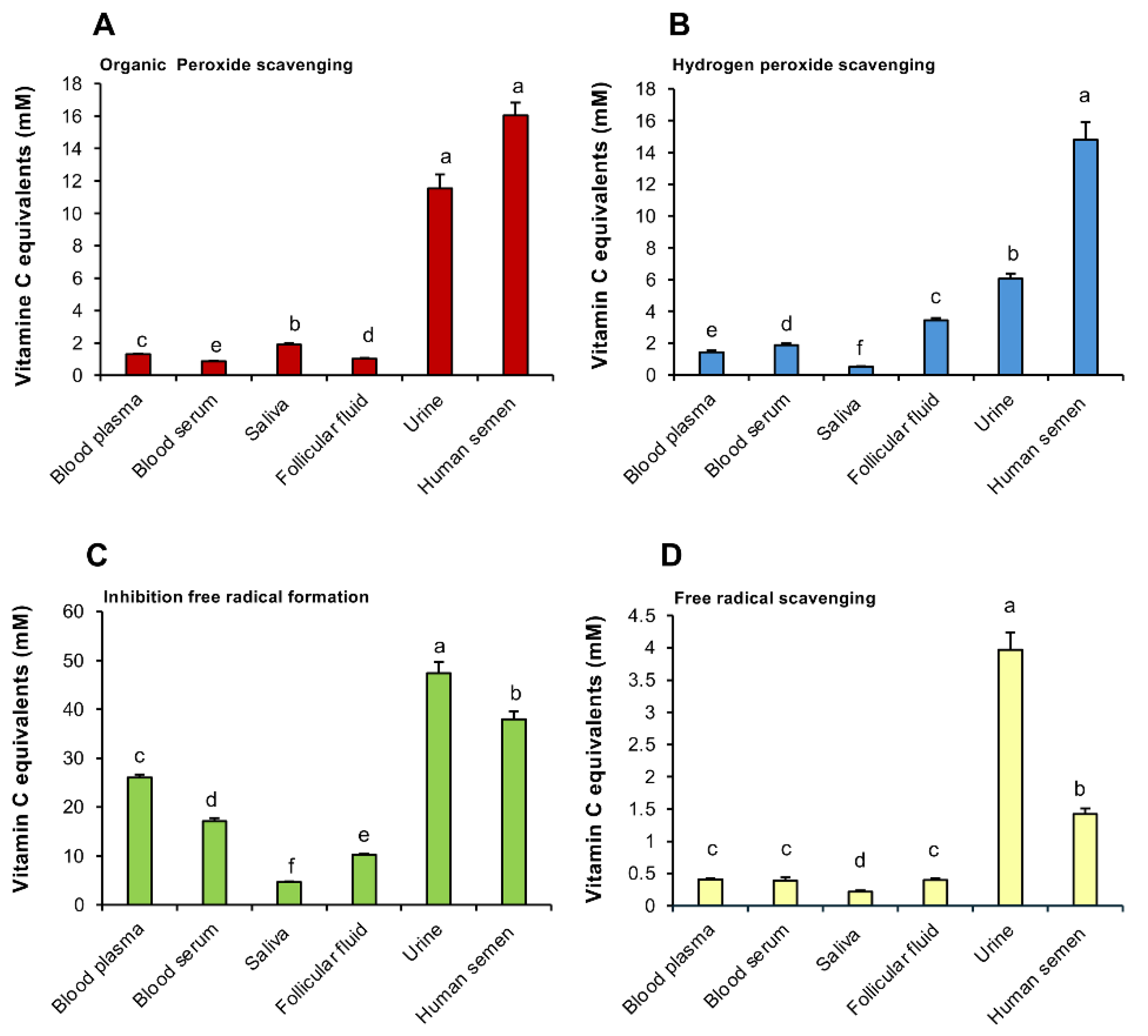
Figure 3.
Antioxidant activity in the seminal plasma in different species. (A) Organic peroxide scavenging activity. (B) Hydrogen peroxide scavenging activity. (C) Inhibition of ABTS•+ radical formation. (D) ABTS•+ radical scavenging activity. All results are expressed as vitamin C equivalents. Statistical analysis by ANOVA and confirmed using non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 9.
Figure 3.
Antioxidant activity in the seminal plasma in different species. (A) Organic peroxide scavenging activity. (B) Hydrogen peroxide scavenging activity. (C) Inhibition of ABTS•+ radical formation. (D) ABTS•+ radical scavenging activity. All results are expressed as vitamin C equivalents. Statistical analysis by ANOVA and confirmed using non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 9.
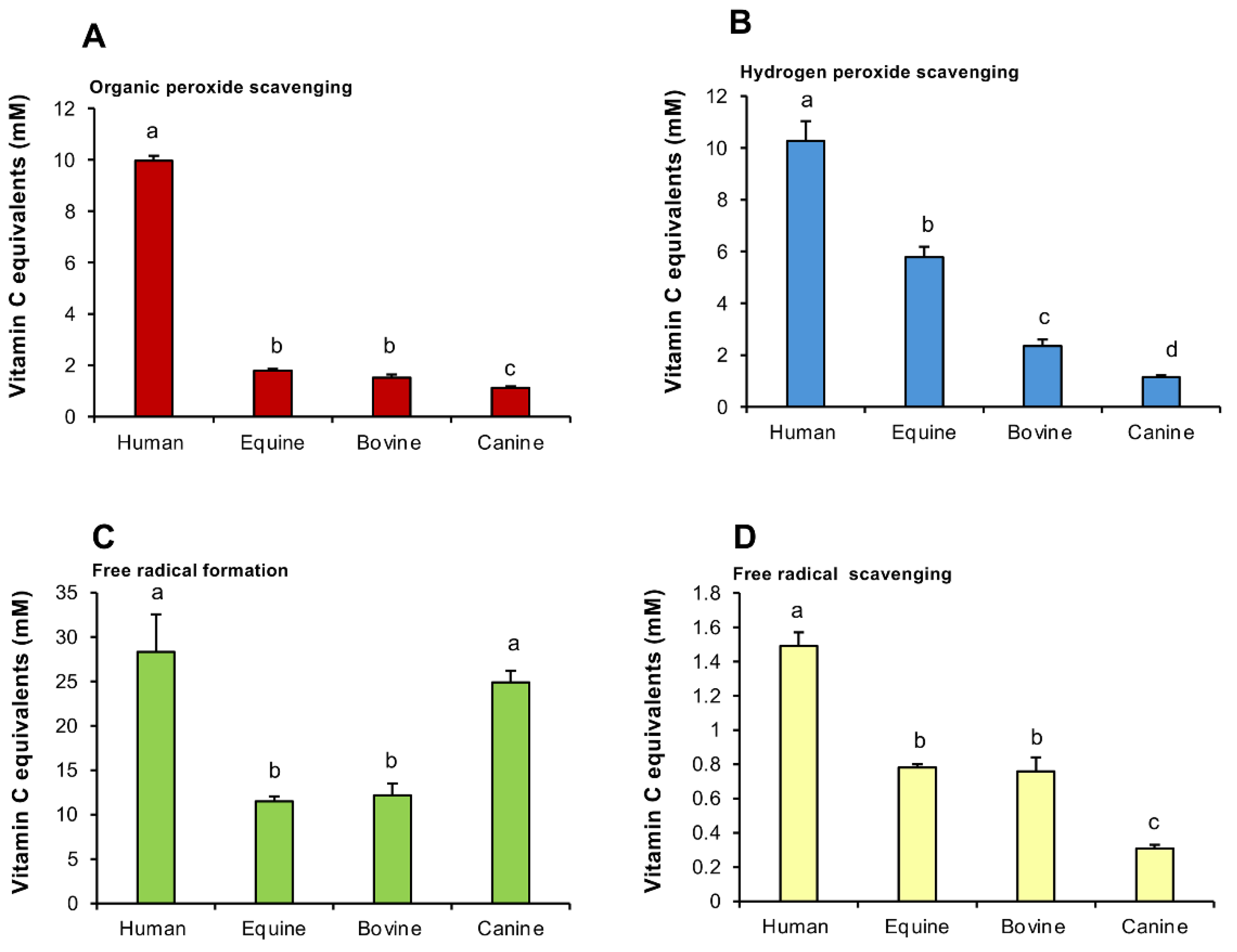
Figure 4.
Dose-dependent impact of cumene hydroperoxide on sperm motility in different species. Both total motility and progressive motility were assessed using a CASA system and two time points were examined: 15-30 min and 2 h. (A) Human spermatozoa after 15-30 min exposure to cumene hydroperoxide. (B) Human spermatozoa after 2 h exposure to cumene hydroperoxide. (C) Bovine spermatozoa after 15-30 min exposure to cumene hydroperoxide. (D) Bovine spermatozoa after 2 h exposure to cumene hydroperoxide. (E) Equine spermatozoa after 15-30 min exposure to cumene hydroperoxide. (F) Equine spermatozoa after 2 h exposure to cumene hydroperoxide. Statistical analysis by ANOVA and confirmed using a non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 3.
Figure 4.
Dose-dependent impact of cumene hydroperoxide on sperm motility in different species. Both total motility and progressive motility were assessed using a CASA system and two time points were examined: 15-30 min and 2 h. (A) Human spermatozoa after 15-30 min exposure to cumene hydroperoxide. (B) Human spermatozoa after 2 h exposure to cumene hydroperoxide. (C) Bovine spermatozoa after 15-30 min exposure to cumene hydroperoxide. (D) Bovine spermatozoa after 2 h exposure to cumene hydroperoxide. (E) Equine spermatozoa after 15-30 min exposure to cumene hydroperoxide. (F) Equine spermatozoa after 2 h exposure to cumene hydroperoxide. Statistical analysis by ANOVA and confirmed using a non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 3.

Figure 5.
Analysis of the impact of human seminal plasma on the cytotoxic impact of cumene hydroperoxide on human spermatozoa. A fixed concentration of purified human spermatozoa (10 × 106/mL) was exposed to 0.25 mM cumene hydroperoxide alone or in the presence of human seminal plasma (3.125 - 12.5%) and the impact on sperm motility assessed using CASA. (A) Total motility (B) Progressive motility. A 15-30 min exposure to cumene hydroperoxide significantly suppressed both motility (p < 0.05) and progressive motility (p < 0.01) however this peroxide -mediated impact was negated by the presence of seminal plasma. Statistical analysis by ANOVA and confirmed using a non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 7.
Figure 5.
Analysis of the impact of human seminal plasma on the cytotoxic impact of cumene hydroperoxide on human spermatozoa. A fixed concentration of purified human spermatozoa (10 × 106/mL) was exposed to 0.25 mM cumene hydroperoxide alone or in the presence of human seminal plasma (3.125 - 12.5%) and the impact on sperm motility assessed using CASA. (A) Total motility (B) Progressive motility. A 15-30 min exposure to cumene hydroperoxide significantly suppressed both motility (p < 0.05) and progressive motility (p < 0.01) however this peroxide -mediated impact was negated by the presence of seminal plasma. Statistical analysis by ANOVA and confirmed using a non-parametric test. All columns not connected by the same letter are significantly different. Data presented as means ± S.E.M.; n = 7.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



