
Submitted:
01 October 2024
Posted:
01 October 2024
You are already at the latest version
Abstract
Research in seaweed cultivation technologies aims to increase production and reduce costs, leading to more efficient and sustainable processes. In this study, we analyzed the outdoor production of Ulva compressa cultured in summertime at different stocking densities of 0.6, 0.8 and 1.0 kg Fresh weight (FW) m-2 in a raceway photobioreactor with 30 m2 surface (3000L) and its relation to pho-tosynthetic activity. Under the experimental conditions of high temperature (> 28-30 oC) and pH > 9 in culture water, higher seaweed density resulted in lower specific growth rate. The biomass pro-duction has been related to photosynthetic activity by using in vivo chlorophyll a fluorescence. Dynamic photoinhibition was observed at noon, which was less severe in cultures with higher algal densities. However, photosynthesis recovered in the afternoon. Seaweeds that were acclimatized for a week to the conditions of 1.0 kg FW m-2 stocking density showed an increase in biomass growth and absence of photoinhibition compared to non-acclimatized thalli. In conclusion, by using high densities (1.0 kg FW m-2), U. compressa presented the highest growth rate in a mid-scale raceway photobioreactor under stressed conditions related to the increase of photosynthetic capacity.

Keywords:
Acclimatization
; Algal culture density
; Photosynthetic activity
; Raceway photobioreactor
; Ulva compressa
; Stress conditions
1. Introduction
Macroalgae provide a multitude of ecosystem services of the following categories, e.g., regulation (carbon fixation, pH increase, biofiltration), supporting (habitat, biodiversity, photosynthesis), cultural (science and education) and supply (food, feed, drugs, fibers, etc.) [1,2]. Indeed, seaweeds are gaining interest in the scientific community, industry and society in general for their potential as a source of valuable, sustainable biomass in the food, feed, chemical and pharmaceutical industries, but also for its interest as biofilters, water quality indicator and other biotechnological applications [3,4,5,6,7].
The aquaculture industry has developed some strategies for cultivating marine organisms, including macroalgae, to increase biomass yields and reduce pollutant release [8,9]. Seaweed aquaculture provides ecosystem services that can improve conditions in coastal waters for the benefit of other living organisms and the environment [10].
The cultivation of seaweeds in open sea systems has advantages compared to land-based system related to productivity, scaling-up and costs. However, sometimes safety issues arise due to the presence of chemical and biological contaminants [11]. Algae biomass coming from harvesting in the natural environment or from aquaculture production at sea could present variability in terms of its quality with respect to the adsorption and absorption of heavy metals from the water that accumulate in the biomass [12,13], especially in areas close to big cities and mining activity.
Land-based aquaculture can also have environmental impacts, particularly if high levels of chemical fertilizers are dumped into the sea, leading to eutrophication. Open systems are more susceptible to environmental contamination from pollutants and unwanted species, while controlling parameters such as temperature and pH can be more challenging. On the other hand, closed systems allow for greater control of parameters such as nutrients and reduce both contamination of the growing medium and the presence of unwanted species, while pH and temperature maintenance is more expensive. These systems offer the possibility of controlling certain production-related traits, such as nutrient concentration, light quantity and quality, and algae concentration [14].
Therefore, solutions based on the cultivation of algae species in land-based systems in which main cultivation parameters are controlled, make this type of systems very interesting for the continuous production of standard-quality biomass to obtain high value biocompounds [15,16]. Species of the genus Ulva have been proposed as a valuable resource for a long time due to their multiple uses and high growth rates [17,18,19]. This genus has been used as efficient biofilters in integrated multitrophic aquaculture (IMTA) systems by using fishpond effluents [20,21]. The importance of this genus has leaded to establish a European Cost Action (CA20106 – Tomorrow’s “wheat of the sea”: Ulva a model for an innovative mariculture) focused on its research [22].
The present work aimed to achieve progress towards the cultivation of the macroalgae Ulva compressa in well-known systems such as raceway ponds, previously assayed in various microalgae species [23,24,25]. This system is based on the closed recirculation of water and nutrients which optimizes the use of these resources as well as the energy consumption.
Specifically, we evaluated the optimal algae density in terms of maximum productivity in this outdoor culture system under non-controlled environmental factors. Lower nutrients input, if compared to other studies [26], were chosen in order to evaluate the ability of this algae to reach optimal productivity with minimum inputs for the sake of a more sustainable and economic culture. Nevertheless, the nutrient content is within the range of previous studies using IMTA systems [27,28].
The experiments were carried out in Southern Spain within the summer period, reaching high temperature in the water (>30 oC) and daily irradiance of photosynthetic active radiation (PAR, λ=400-700 nm) higher than 12000 kJ m-2 [29]. The ability of Ulva to live and grow above this water temperature has been previously demonstrated [30,31], but there is lack of evidence about the effect of the algae density on the physiology and productivity in this culture system, especially the effect of high both dissolved oxygen and pH due to high photosynthetic capacity. It is known that Ulva spp. can survive in supralittoral ponds under high temperature and pH and increasing the salinity due to water evaporation, and also can grow under low salinity [32,33]. Ulva spp. present highly efficient different enzymatic systems for bicarbonate assimilation under high pH [34,35].
Once the optimal thalli density was determined, we also analysed the effect of acclimatization to these environmental conditions. Growth rate is related to the photosynthetic activity estimated by using in vivo chlorophyll a fluorescence of photosystem II (PAM fluorometers)[36,37]. This technique has been proven as good tool in seaweeds to evaluate both photosynthetic efficiency and capacity through maximal quantum yield (Fv/Fm) as an indicator of photoinhibition and Electron Transport Rate (ETR) as an estimator of photosynthetic capacity (gross photosynthesis), respectively [36,37,38]. Good correlation between gross photosynthesis and ETR has been found both in green micro and macroalgae [27,36,39,40]. In situ monitoring of effective quantum yield under solar radiation allowed to determined daily productivity under different environmental conditions in Ulva spp. growing in tanks [27,39,41].
2. Results
2.1. Diurnal Variation in Water Physical and Chemical Variables during the Course of the Culture.
The correspondence of pH with the rest of variables is shown in Figure 1.
The pH and DO increased from the morning to midday reaching the maximum values close before noon (Figure 1A). This pattern must be related to a positive net photosynthesis. Depending on the biomass density, different patterns are observed on day 4. At the lowest density (0.6 kg FW m-2) and the highest (1.0 kg FW m-2) a drop in pH values was observed, which implies a reduction in net photosynthesis; while in the density of 0.8 kg FW m-2 an increase in pH was observed on day 4, which would mean an increase in the net photosynthesis of the system, but a decrease was observed later.
Around midday, a sharp decrease in pH and DO was found at temperatures around 27ºC (Figure 1B). This indicates that high irradiance induces a reduction of Electron transport rate as estimator of gross photosynthesis possibly due to photoinhibition (Figure 1C), whereas high temperatures increase respiration in addition reducing oxygen solubility in water. The drop in DO at noon was produced after the maximal irradiance and coupled to the maximal temperature. Interestingly, maximal temperature was produced 1-2 h after the maximal irradiance (Figure 1), then in the early afternoon recovery of DO was produced. A small, transitory increase in pH was observed (bold arrows in Figure 1A), that imply a positive net photosynthesis. This transitory peak in photosynthesis induces a reduction of the negative slope in DO pattern.
2.2. Biomass Growth Rate and Nutrient Assimilation in the Raceway Ponds.
For 0.6, 0.8 and 1.0 kg FW m-2 densities, biomass growth rate values at FW basis were 36.7, 39.1, 38.1 g FW m-2 day-1, whereas at DW basis were 6.23, 6.64, 6.48 g DW m-2 day-1, respectively (Table 1).
SGR of 5.94, 4.90, 4.07 % day-1 were found for mentioned culture densities in our study (Table 1). Biomass FW increments were 7.7, 8.2 and 8.3 kg FW week-1 for the 0.6, 0.8 and 1.0 kg FW m-2 densities, respectively (Table 1).
When normalizing biomass increment to nutrient consumption (i.e., yield per N and P), 0.611, 0.651 and 0.659 kg FW biomass mg-1 N, 8.280, 8.817 and 8.925 kg FW biomass mg-1 P was produced, as can be observed in Table 1 at 0.6, 0.8 and 1.0 kg FW m-2 biomass concentration respectively.
Regarding nutrient assimilation, 900 µmol L-1 of nitrogen and 54 µmol L-1 of phosphate was injected weekly in the culture and the nutrient concentration always was depleted in the water, i.e., the NUE reached values of 100% in every algal density. NUR values were 5.952, 4.464 and 3.571 µmol N g DW-1 h-1 in 0.6, 0.8 and 1.0 kg FW m-2, respectively.
2.3. Effect of Algal Density on Physiological and Functional Variables.
2.3.1. In Situ Photosynthetic Activity.
As shown in Figure 2A, there was a gradual rise in the value of YII except 24 hours after starting the experiment when a pronounced fall was observed. This indicates a stress of the algae since the algae were transferred from tanks at density of 30 g FW L-1, i.e., from shade environment in the tanks to a raceway with higher solar radiation exposure. As in the case of DO (Figure 1A), a decrease of ETR values were observed at noon under maximal irradiances and a subsequent recovery of normal values. The in situ electron transport rate (ETRin situ) (Figure 2B) presented a similar pattern to that observed in YII (Figure 2A), with a pronounced drop in values within 24 hours from the beginning of the experiment in all densities assayed.
2.3.2. Ex Situ Photosynthetic Activity: Rapid Light Curves (RLC).
Maximal quantum yield (Fv/Fm) was higher under the highest algal density than that under 0.6-0.8 kg FW m-2 (Table 2).
The photosynthetic efficiency (αETR) was not significantly different among the different algal densities. However, the saturation irradiance (Ek) increased in cultures under 1.0 kg FW m-2 density in spite of the self-shading.
Maximal non photochemical quenching (NPQmax) was higher under 1.0 kg FW m-2 density compared to lower algal densities, while ETRmax/NPQmax decreased under 1.0 kg FW m-2 algal density cultures compared to 0.6 kg FW m-2.
Maximal ETR under outdoor raceway ponds (ETRin situ) reached values of 150-160 µmol electrons m-2 s-1, whereas ex situ ETRmax reached values of 90-94 µmol electrons m-2 s-1.
2.4. Effect of Algal Acclimatization on Physiological and Functional Variables.
Acclimatized algae coming from 1.0 kg FW m-2 stocking density (as explained in section 4.2) showed an improvement in growth related parameters (Table 3). SGR and biomass increment raised in 0.7% day-1 and 1.7 kg FW week-1, respectively. This biomass increment corresponds to a 20.8% in 1 week compared with the non-acclimatized algae. Improvements in the efficiency in nitrogen and phosphorus assimilation were also found, from 0.66 to 0.79 kg FW mg-1 N and 8.93 to 10.75 kg FW mg-1 P.
Photosynthetic activity, expressed as in situ YII and ETR, were higher in acclimatized- than that in non-acclimatized algae (Figure 3). Both photosynthetic parameters improved, specially between 8h and 32h after the beginning of the culture.
All photosynthetic parameters determined by in vivo Chl a fluorescence increased in acclimatized algae (Table 4). The algae present a metabolism of sun type algae (increased Ek and ETRmax), revealing its higher photosynthetic capacity. The NPQmax decreased in acclimatized algae. On the other hand, as expected the ETRmax/NPQmax increased.
Many research studies on Ulva spp. production have been conducted, as shown in Table 5 (modified from [26]). Different algal densities, variety of nutrient source, different tank volumes, and number of water exchanges have been studied. Our study makes use of chemical fertilizers under low concentration compared to other works and with no water exchanges. Table 5 shows the great variability between the data obtained in growth rate by different authors, included in this study. This is due to the diversity of tanks, volumes, etc. used in each study. A comparison of biomass productivity expressed as g DW m-2 day-1, reveals significant discrepancies between studies. However, when this data is converted into g L-1 day-1, the differences are less pronounced.
2.5. Functional Relationship between Variables.
Values of Pearson correlation between different functional variables are shown in Table 6. A correlation was observed between SGR and nitrate uptake rate (NUR) as well as between NUR and the ETRmax/NPQmax ratio. However, no correlation was found between SGR or NUR and ETRmax.
3. Discussion
3.1. Physical and Chemical Variables during the Culture.
pH, DO and Temperature presented variations depending on the concentration of biomass of the reactor and the time of the day (Figure 1). The phenomenon of dynamic photoinhibition, characterised by a decline in pH and dissolved oxygen (DO) levels, followed by a subsequent recovery, has been observed in contrast to the more prolonged and irreversible process of chronic photoinhibition, which occurs without a return to normal levels. Dynamic photoinhibition is regarded as a physiological strategy of photoprotection. While it does affect productivity, it allows for the reduction of radical oxidative substances, thereby limiting the damage to pigments [52,53,54].
In the afternoon, with the decrease in incident solar radiation (between 1200 – 1300 µmol photons m-2s-1), Ulva compressa can use bicarbonate due to high activity of carbonic anhydrases [34]. Nevertheless, it has been demonstrated that CO2 addition improved photosynthetic activity and increased productivity of Ulva, provided that pH does not rise above 11-12 [55]. In this study, in the periods with high pH (>9) (Figure 1 A, B, C), bicarbonate must be the main source of carbon, and it is transformed to CO2 by the action of carbonic anhydrase enzymes. Thus, the efficient use of bicarbonate is an advantage because it reduces the costs (since the expense of CO2 is one of the limitations in many algal cultures), avoiding risks of contamination by other algae that cannot grow at such high pH [34]. Limitations in access to carbon (in the form of CO2 or bicarbonate) cause a substantial reduction in the photosynthesis of the algae and reduce its productivity, giving rise to a negative net photosynthesis and a prevailing respiratory process [56]. Similarly, we found that both pH and DO in the raceway decrease even during the hours of high radiation, although later at sunset the pH is temporarily recovered to fall again later during night (Figure 1A).
3.2. Biomass Growth Rate and Nutrient Assimilation in the Raceway Ponds.
The SGR decrease as the density of our culture increases (Table 1). Similar results were obtained in other studies (6,16,26), where tanks were used as culture systems at higher algal density than in our work (1.0 – 3.0 kg FW m-2). [17] conducted a screening study on 48 strains of Ulva compressa. and found an SGR average between 3 - 6% day-1, being U. pseudorotundata one of the species with the highest SGR together with Ulva prolifera. Considering the night period, the average SGR was 8.63% but certain Ulva strains can reach specific growth rate of 12 - 16% per period-1 [17]. The diversity in the amino acid content among the strains was also very high. Thus, it is crucial to investigate the response of different strains in terms of growth and biochemical contents at similar culture conditions in order to optimize the productivity of high value biocompounds, as it has been proven in diverse screening studies mainly for food and cosmeceutical applications [57,58,59,60].
In contrast with SGR, biomass FW increased at higher initial densities, ranging from 7.7 to 8.3 kg FW week-1. This is not a remarkable increase of production, but under the highest initial biomass of Ulva, less competition with microalgae in the reactor was visually observed compared to lower initial densities. Therefore, the higher the initial density, the less competition for nutrients was taking place. When normalizing biomass increment to nutrient consumption i.e., yield per N, cultures with higher density presented higher productivity yield, indicating a more efficient use of nutrients, i.e., more biomass was produced with less nutrient consumption (Table 1).
As mentioned above, algal density showed an inverse correlation with SGR, but the highest density presented the highest total production under our experimental conditions. In the present work, a low concentration of nutrients was used compared to other studies, i.e., a total of 0.06 mg N m-2 day-1 of nitrogen input, much lower than in other studies (e.g., [26] used 0.81-0.99 g N m-2 day-1). Still, these nutrient concentrations are much higher than that in the coastal Mediterranean waters [61]. In any case, the content of inorganic nitrogen used in this study is in the range of the nitrogen levels in the fishpond effluents under IMTA (27,28,62,63). Thus, if increasing nutrient input, an increase in biomass productivity could be expected.
Values of NUE and NUR found in our work were similar to other studies [5] (NUE 100% in every algal density, NUR values of 5.952, 4.464 and 3.571 µmol N g DW-1 h-1 in 0.6, 0.8 and 1.0 kg FW m-2, respectively).
3.3. Effect of Algal Density on Physiological and Functional Variables.
As in the case of DO (Figure 1A), a decrease of ETR values were observed at noon under maximal irradiances. Thus, if we consider ETR as an estimator of photosynthetic capacity and biomass productivity [39,40], this result indicates a decay of the biomass productivity due to photoinhibition by excess of oxygen i.e. production of radical oxygen substances (ROS). However, this transient response since recovery of ETR values were observed in the afternoon. Recovery at different times of the day was observed as a consequence of the algae acclimatization to the new culture conditions. Under the highest culture density, the lowest photoinhibition in the central hours of day (5, 29 and 149 hours) is found as a consequence of the self-shading and subsequent photoprotection pattern, as reported by [52].
A pronounced drop in ETRin situ values within 24 hours from the beginning of the experiment in all densities assayed is observed (Figure 2). This decrease could be related to stress due to excess of radiation, as obtained in other studies for Ulva under different physiological conditions [27,41].
The positive correlation between in situ maximal quantum yield (Fv/Fm) and algal density (Table 2) confirmed photoprotection against photoinhibition by self-shading of the thalli in the ponds, as reported in [52]. The ETRmax was similar among the different algal densities, reaching values higher than that reported in U. pseudorotundata growing in tanks moving with an air pump and fed with fishpond effluents [6].
The ex situ photosynthetic activity revealed that, under the three algal densities assayed, the Ek values corresponded to sun-type algae, as it has been reported in other Ulva species growing in tanks [27,64]. Sun-type algae (i.e., intertidal algae) presented higher ETR and Ek and lower αETR than algae from shade habitats (subtidal algae) [36,65].
High NPQ (Table 2) indicates high capacity for energy dissipation. This photosynthetic parameter is an indicator of optimal photoprotection, since NPQ is the ratio between two yield losses, namely, YNPQ and YNO, being YNPQ a mechanism related to the dissipation of energy as heat and fluorescence through photoregulated mechanisms (i.e., xanthophyll cycle) and YNO is passive dissipation [66]. Higher YNPQ than YNO is an indicator of optimal photoprotection under high irradiance or other stress on photosynthesis as high temperature or increased UV radiation [67,68]. The ETRmax/NPQmax ratio has been used as a physiological indicator expecting to be higher under optimal growth conditions, namely, when production is greater than energy dissipation. Conversely, this ratio is anticipated to decrease under increased UV radiation [67] or under acidification and low nitrogen conditions [27]. However, in this study the ETRmax/NPQmax decreased under 1.0 kg FW m-2 algal density cultures compared to 0.6 kg FW m-2. Under 1.0 kg FW m-2 it seems like energy dissipation or yield losses are higher to acclimatize to the environmental conditions, i.e., nutrient competition under high solar exposure. Nevertheless, despite the comparable ETRmax and diminished ETRmax/NPQmax in 1.0 kg FW m-2 relative to the 0.6 kg FW m-2 alga density, the biomass yield remained consistently high.
Higher values of in situ photosynthetic rate compared to ex situ, both by fluorescence determination (ETR) and oxygen evolution (gross photosynthesis) has been previously reported in Ulva spp. [69] and other micro and macroalgal species [39,40]. High correlation between ETR and gross photosynthesis has been reported in Ulva species and thus ETR has been demonstrated to be a good indicator of gross photosynthesis (27,36,70). The highest in situ ETR compared to ex situ has been explained due to the incident actinic light. Under in situ condition, algae are exposed to solar radiation and under ex situ measurements the actinic light presents a narrow spectrum (red light), thus a broader range of quanta are available to conduct photosynthesis under in situ compared to ex situ [27,39]. In addition, under laboratory conditions, a unique piece of thallus is used to conduct the RLC and although three replicates are conducted, each thallus was continuously exposed to increasing irradiance for several minutes. Under these conditions, acclimatization to light can be performed and consequently YII can be reduced. In contrast, under ex situ measurements, eight different thalli randomly selected in the culture were used, and there is no exposure to increasing irradiances as when performing the RLC [27,39].
3.4. Effect of Algal Acclimatization on Physiological and Functional Variables.
The improvement in growth parameters observed (Table 3) is due to an increase in photosynthetic capacity (Figure 3) by acclimatization to the stressful processes of the culture medium, such as temperature, light intensity, as well as nutrient deficiency in the culture medium. The afternoon recovery i.e increase of YII (from 5h to 8h, corresponding to 2.00 pm and 5.00 pm, respectively) was higher in acclimatized algae, which contributed to higher ETRin situ, indicating that more energy can be available for growth [37].
As we explained in point 2.4, the algae presented a sun-type metabolism (high Ek and ETRmax). The reduction of NPQmax indicates that the algae require less energy dissipation because of physiological acclimatization to the high irradiance. The increment of ETRmax/NPQmax indicates that algae have more energy available for growth as it has been previously reported under optimal growth conditions in other seaweeds including Ulva spp. (27,39,67).
3.5. Functional Relationship between Variables.
SGR correlated to nitrate uptake rate (NUR) since photosynthetic productivity is related to the availability and assimilation rate of nitrate [39]. NUR correlated to ETRmax/NPQmax but not with ETRmax, indicating that the nitrate assimilation rate and specific growth is more related to the ratio between productivity and energy dissipation than with only productivity. If the growth is represented as specific growth rate in % d-1, a positive correlation with ETRmax/NPQmax was found, i.e., highest SGR and ETRmax/NPQmax at 0.6 kg FW m-2 (see Table 6).
The data shown in this study have been obtained in the month period with the highest irradiance and temperature of the year in Malaga, demonstrating that U. compressa can be cultivated in this type of photobioreactors under high solar exposure. Under these conditions, more energy is available for photosynthesis but also, under very high temperature, there is an increase in respiration rate and consequently gross photosynthesis is expected to be increased and more ATP is produced for the assimilation process. Good correlation between gross photosynthesis measured by oxygen evolution and electron transport rate has been found in Ulva spp. [36,70]. High respiration can provide additional ATP to maintain both inorganic carbon and nitrogen assimilation and, in addition, to contribute to photoprotection. On the other hand, high oxygen consumption due to Mehler reaction is expected under the environmental condition in this study [36]. This capacity to live and grow at water temperatures above 30 oC of different Ulva strains [30,31], implies its potential as a future cultivated algae for multiple uses in a perspective of climate change in which higher temperatures are expected in southern European countries, such as Spain. In previous works, an induced increase in temperature of 4°C, which represent a pessimistic scenario for climate change at the end of this century, was found to result in an increase in both gross photosynthesis and ETRmax in Ulva rigida [27]. However, the observed increase in both photosynthetic parameters was produced only under high nitrate conditions, at both low and high carbon levels. Conversely, the decrease was produced under low nitrogen and high CO2 conditions (acidification). Therefore, it can be concluded that the adaptation of Ulva to climate change is not solely dependent on temperature, but also on the levels of nitrate and CO2 present in the water.
4. Materials and Methods
4.1. Sampling
The green alga Ulva compressa (also called mutabilis), (Linnaeus, 1753) used in this study was collected in salt marsh areas of the bay of Cádiz (36°30'N, 6°10'W). The algae were transported in thermal boxes to the Grice Hutchinson Research Centre at Malaga University where the experiments were conducted.
Before the establishment of the culture in raceway systems, thalli of Ulva compressa were maintained at stocking densities of 30-50 g L-1 outdoor in 300 L square tanks with continuous aeration, in reactors with artificial seawater (35 psu) and supplemented 3 times every week with NH4NO3 and KH2PO4 as described below.
4.2. Experimental Conditions
Experiments were conducted during June-July 2022. For the evaluation of algae density, stocked thalli were transferred from the stocking tanks to a raceway pond of 30m2 surface area and 10 cm depth, i.e., a volume of 3000 L. Concentrations of 150 µmol L-1 NH4NO3 and 18 µmol L-1 KH2PO4 were added to the reactor 3 times per week by means of agricultural granular fertilizers of the brand Fertiberia® (34.5% N purity) and Fenasa® (22.7% P purity), respectively. These nutrient values are in the range used in IMTA systems in other works (34,70,71). The algae were grown unattached and kept in movement by the paddlewheel of the raceway reactor. A continuous aeration in the sump was blown without addition of CO2.
As explained in Figure 4 , thalli were grown for 7 days in each treatment, which consisted of different thalli densities (0.6, 0.8 and 1.0 kg FW m-2). In all treatments, the input biomass came from the same 300 L stocking tanks. Densities were tested sequentially using the same raceway system, i.e. first week at 0.6 kg FW m-2, second week at 0.8 kg FW m-2 and third week at 1.0 kg FW m-2. At the end of each treatment (day 7), the algae were completely collected and growth variables (specific growth rate, biomass increase and N consumption) were measured. The new density treatment was then set up and the same procedure was carried out.
Physico-chemical parameters of the culture (pH, dissolved oxygen (DO) and water temperature, as well as incident irradiance) were monitored throughout the experiments in each treatment (see section 2.3), as well as physiological and functional variables (photosynthesis as in vivo chlorophyll a fluorescence - related parameters, as described in section 2.4).
In order to evaluate physiological changes and growth patterns after acclimatisation, a further experiment was carried out at 1.0 kg FW m-2 density, algae tested at this density were collected from the raceway reactor at the end of the treatment (1 week) and re-established at the same initial density of 1.0 kg FW m-2 after measuring growth-related variables. The culture was then continued for an additional week, after which physiological and growth-related parameters were measured using the same procedure as in the previous test. For the post-acclimatisation treatment, the input biomass from the pre-acclimatised 1.0 kg FW m-2 density collected at the end of this treatment was used (Figure 4).
4.3. Water Physical and Chemical Analysis.
Water temperature (T), pH and DO were monitored automatically every 30 min along the experiments using raceway-integrated sensors (Jumo tecLine HD, Germany). Irradiance was measured automatically every 30 minutes outside the tank as incident irradiance, using HOBO pendant® Light Data Logger (Onset, MA, USA). In order to verify the automatic measurements, water T, pH, salinity and DO were measured 3 times every day manually using portable devices (LAQUAact pH, EC120 and HI S8193 Dissolved Oxygen, respectively) at 9:00, 12:00 and 15:00.
To determine changes in concentration of N-NH4+, N-NO3- and PO4 3-, water samples were taken before addition of nutrients and then it was analysed colorimetrically using a continuous flow automated analyser (Technicon AA-2), following the procedure of Grasshoff et al. [1983].
The reduction in nitrogen concentration between the time intervals is expressed as percentage and defined as “nutrient uptake efficiency” (NUE) and was calculated by Eq. 1 assessing the changes in total nitrogen concentration:
NUE (%) = 100-[Ct+1*100/Ct] (eq. 1)
Where Ct represents the initial concentration of nutrients and Ct+1 represents the concentration after t+1.
The amount of nitrogen removed per unit of time per volume by seaweed dry weight represent the “nutrient uptake rate” (NUR) and is determined from changes in nitrogen, according to Eq. 2:
NUR (µmol N g-1 DWh-1) = [(Ct*Vt) - (Ct+1*Vt+1)]/(B*Δt) (eq. 2)
Where Ct represents the initial concentration of nitrogen, Vt represents the initial volume of the photobioreactor (in L), Ct+1 represent the concentration of nitrogen after t+1, Vt+1 represent the volume of the photobioreactor after t+1 (in L); B represents dry biomass used initially (g), and Δt represents the time interval between t and t+1 in hours.
4.4. Biomass Growth Parameters and Physiological Variables Measurements.
For every assayed density, growth parameters were determined at the end of every experiment (as explained in 2.2) by measuring the increment in algal fresh biomass after harvesting. Thalli were collected with a net and manually pressed always by the same person in order to drain out the excess water.
Biomass growth rate parameters were calculated as the biomass increment in fresh weight (FW) or dry weight (DW) basis normalized to units of volume or area. The Specific Growth Rate (SGR) was calculated following the equation:
SGR= 100 x [ln (Wf/W0)] / t (eq. 3)
where W0= initial biomass, Wf = final biomass, and t expresses the days of culture in the experimental set (adapted from [79]).
Relative biomass increment was measured following the equation:
Relative biomass increment= (Wf-W0)/Wf (eq. 4)
where W0= initial biomass, Wf = final biomass
Photosynthetic activity was estimated through the in vivo chlorophyll a fluorescence associated to photosystem II (PSII) by using a Mini-PAM-II fluorometer (Walz GmbH, Effeltrich, Germany), with red light as measuring, actinic and saturating pulse light for both in situ in the raceway ponds and ex situ in the laboratory measurements.
(a) In situ – effective quantum yield (YII) (eq. 5) was determined outdoor in algae growing in the raceway pond. YII was used for the determination of Electron Transport Rate (ETR) (eq. 6) as estimator of photosynthetic capacity (gross photosynthesis) and algal productivity [40]. YII was determined as follows: firstly, fluorescence at steady state (Ft) was calculated by measuring red light and then saturating light pulse was applied (800 ms, 5000 µmol m-2 s-1) to algal thalli in the reactor to determine maximal fluorescence at light-acclimated samples (F’m), according to [27]. Effective quantum yield was calculated as:
YII= (F’m - Ft) / F’m (eq. 5)
ETRin situ was calculated as:
ETRin situ= YII x EPAR x A x FII (eq. 6)
Where EPAR is the irradiance of photosynthetic active radiation (λ=400-700 nm) of the incident light at the surface of the reactor; A is the absorptance, which was measured every day according to [68]; FII is the fraction of chlorophyll a associated to PSII being 0.5 in green algae [67].
YII and ETRin situ were quantified at different hours of local time (9:00, 14:00 and 17:00) and in different days from 8 random algal samples of the reactor.
(b) Ex situ – For rapid light curve measurements, 3 replicates were taken from raceway ponds, transported to the laboratory and introduced in 50 mL tubes covered by dark foil. Firstly, algae were incubated 15 min darkness and basal fluorescence was determined by switching on measured light (Fo), and then saturation light pulse was applied to measure maximal Florescence (Fm). Maximal quantum yield (Fv/Fm) was determined as:
Fv / Fm= Fm - Fo / Fm (eq. 7)
Then algal samples were exposed for 30 s to twelve increasing irradiances (25, 45, 66, 90, 125, 190, 285, 420, 625, 845,1150, and 1500 μmol photons m−2 s−1) of actinic red light followed by a saturating light pulse determining effective quantum yield (YII) and ETR as it is indicated above (equations 5 and 6). This ETR is denominated ex situ ETR. ETR versus irradiance obtained from light curves were fitted according to [71] models to estimate the variables of maximal electron transport rate, i.e., ETRmax, photosynthetic efficiency (⍺ETR) and saturated irradiance (Ek).
The non-photochemical quenching of fluorescence (NPQ) is calculated as follows:
NPQ = (Fm – F’m) / F’m (eq. 8)
NPQ max is calculated by the fitting of NPQ versus irradiance function by using [71] model.
NPQ can also be expressed as the ratio YNPQ/YNO according to [66]. YNO is the fraction of energy passively dissipated as heat and fluorescence, mainly due to closed PSII reaction centres. High values indicate an inability of the macroalga to protect itself against photodamage by an excess of radiation. YNPQ is the fraction of energy dissipated as heat via regulated photoprotective mechanisms. High values are indicative of photoprotective capacity. The ratio ETRmax/NPQmax represents an estimator of the ratio between productivity and energy dissipation, according to [67].
4.5. Statistical Analysis
Statistical analyses were performed using Statgraphics Centurion 19.3.02 software. One-way ANOVAs were performed for photosynthetic variables in both trials (density and acclimatization) after verification of the fulfilment for data normal distribution and homogeneity of variance. Otherwise, a non-parametric Kruskal-Wallis analysis were done. For means separation after ANOVAs, Tukey’s test was used. Pearson correlation between response variables was performed using Statistica 7 software.
5. Conclusions and Future Applications
As observed in this study, raceway pond photobioreactors can be used for biomass production of Ulva compressa in recirculating conditions. The increase of biomass productivity at the highest algal density was related to the increase of photosynthetic capacity.
Optimal biomass productivity was 0.32-0.36 g FW L-1 day-1 (6-8 g DW m-2 day-1). These values are in the range of microalgal biomass productivity in this type of photobioreactor [72], by agriculture fertilizers were added at very lower amounts than in other studies [26], simulating the nutrient range reached by using fishpond effluents (62,63,73). In spite of this, we consider that there is a limitation of nutrients in our culture and the increase of nutrients would allow us to increase biomass production. The best concentration of biomass for culture in circumstances similar to ours could be 0.8-1.0 kg FW m-2, since they produce same or even higher amount of biomass and can compete better with the microalgae that may appear in the reactor when it is open air and water is recirculated continuously. It could probably improve if the amount of water is increased in the raceway pond to 20cm depth (200 L m-2) or even 30cm (300 L m-2) and the nutrient concentration is increased, which would result in a higher self-shading, less temperature variability along the day and nutrient availability.
Ulva biomass produced in race ways could be used to extract bioactive compounds and is suitable for use in feed due to its good protein content (13.7-38.9% protein) (5,62,74,75), which allows this industry to be more sustainable and resilient as the market demands [76,77,78]. This also applies to the manufacture of biofertilizers and biostimulants using algae as raw material [79,80,81]. In addition, Ulva in raceway systems due to the high nutrient consumption can supply an ecosystem service related to nutrient biofiltration. Ulva strains have been used for bioremediation and water purification system for both industry, farms and urbans [82,83,84] and it can reach similar biofiltration capacity as microalgae [85,86,87] but with cheaper harvesting process since it is not necessary flocculation or centrifugation, making this species a source of low-cost compounds. In addition, aquaculture of high productive Ulva strains could contribute to the mitigation of climate change although there is a controversy on the actual contribution of carbon sink and sequestration [88,89].
More biotechnological research is necessary to get sustainable cultures of Ulva strains reducing the cost in the frame of Blue economy.
Author Contributions
Conceptualization, Victor Robles-Carnero; Data curation, Victor Robles-Carnero, Rafael Sesmero and Felix L.Figueroa; Formal analysis, Victor Robles-Carnero, Rafael Sesmero and Felix L.Figueroa.; Funding acquisition: Felix L.Figueroa; Investigation, Victor Robles-Carnero, Rafael Sesmero and Felix L.Figueroa; Methodology, Victor Robles-Carnero, Rafael Sesmero and Felix L.Figueroa; Project administration, Felix L.Figueroa; Resources, Rafael Sesmero and Felix L.Figueroa; Supervision, Rafael Sesmero and Felix L.Figueroa; Validation, Victor Robles-Carnero, Rafael Sesmero and Felix L.Figueroa; Writing – original draft, Victor Robles-Carnero, Rafael Sesmero and Felix L.Figueroa; Writing – review & editing, Rafael Sesmero and Felix L.Figueroa. All authors contributed to the article and approved the submitted version.
Funding
This research was financed by the Ministry of Science and Innovation of Spain Government (Alga Hub Project TED2021-131555B-C22).
Acknowledgments
We are also grateful to Dra. Paula Celis-Plá for data curation and reviewing.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bermejo R, Buschmann A, Capuzzo E, Cottier-Cook E, Fricke A, Hernández I, et al. State of knowledge regarding the potential of macroalgae cultivation in providing climate-related and other ecosystem services: a report of the Eklipse Expert Working Group on Macroalgae cultivation and Ecosystem Services. 76 p.
- Duarte CM, Bruhn A, Krause-Jensen D. A seaweed aquaculture imperative to meet global sustainability targets. Vol. 5, Nature Sustainability. Nature Research; 2022. p. 185–93. [CrossRef]
- Charrier B, Abreu MH, Araujo R, Bruhn A, Coates JC, De Clerck O, et al. Furthering knowledge of seaweed growth and development to facilitate sustainable aquaculture. Vol. 216, New Phytologist. Blackwell Publishing Ltd; 2017. p. 967–75. [CrossRef]
- Jusadi D, Ekasari J, Suprayudi MA, Setiawati M, Fauzi IA. Potential of Underutilized Marine Organisms for Aquaculture Feeds. Front Mar Sci. 2021 Feb 11; 7:1250. [CrossRef]
- Massocato T, Robles-Carnero V, Vega J, Bastos E, Avilés A, Bonomi-Baru J, et al. Short-term nutrient removal eficiency by Ulva pseudorotundata (Chlorophyta): potential use for Integrated Multi-Trophic Aquaculture (IMTA). 2022. [CrossRef]
- Massocato TF, Robles-Carnero V, Moreira BR, Castro-Varela P, Pinheiro-Silva L, Oliveira W da S, et al. Growth, biofiltration and photosynthetic performance of Ulva spp. cultivated in fishpond effluents: An outdoor study. Front Mar Sci. 2022 Sep 2; 9:1550. [CrossRef]
- Vega J, Schneider G, Moreira BR, Herrera C, Bonomi-Barufi J, Figueroa FL. Mycosporine-Like Amino Acids from Red Macroalgae: UV-Photoprotectors with Potential Cosmeceutical Applications. Applied Sciences 2021, Vol 11, Page 5112. 2021 May 31;11[11]:5112. [CrossRef]
- Neori A, Chopin T, Troell M, Buschmann AH, Kraemer GP, Halling C, et al. Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofiltration in modern mariculture. Aquaculture. 2004 Mar 5;231[1–4]:361–91. [CrossRef]
- Buschmann AH, Camus C, Infante J, Neori A, Israel Á, Hernández-González MC, et al. Seaweed production: overview of the global state of exploitation, farming and emerging research activity. Eur J Phycol. 2017 Oct 2;52[4]:391–406. [CrossRef]
- Kim JK, Yarish C, Hwang EK, Park M, Kim Y, Kim JK, et al. Seaweed aquaculture: cultivation technologies, challenges and its ecosystem services. Algae. 2017 Mar 15 ;32(1):1–13. [CrossRef]
- Monti M, Minocci M, Beran A, Iveša L. First record of Ostreopsis cfr. ovata on macroalgae in the Northern Adriatic Sea. Mar Pollut Bull. 2007 May 1;54(5):598–601. [CrossRef]
- A Akcali I, Kucuksezgin F. A biomonitoring study: Heavy metals in macroalgae from eastern Aegean coastal areas. Mar Pollut Bull. 2011 Mar 1;62(3):637–45. [CrossRef]
- B Baumann HA, Morrison L, Stengel DB. Metal accumulation and toxicity measured by PAM—Chlorophyll fluorescence in seven species of marine macroalgae. Ecotoxicol Environ Saf. 2009 May 1;72(4):1063–75. [CrossRef]
- García-Poza S, Leandro A, Cotas C, Cotas J, Marques JC, Pereira L, et al. The Evolution Road of Seaweed Aquaculture: Cultivation Technologies and the Industry 4.0. International Journal of Environmental Research and Public Health 2020, Vol 17, Page 6528. 2020 Sep 8;17(18):6528. [CrossRef]
- Dini I. The Potential of Algae in the Nutricosmetic Sector. Molecules 2023, Vol 28, Page 4032 . 2023 May 11 ;28(10):4032. [CrossRef]
- Zertuche-González JA, Sandoval-Gil JM, Rangel-Mendoza LK, Gálvez-Palazuelos AI, Guzmán-Calderón JM, Yarish C. Seasonal and interannual production of sea lettuce (Ulva sp.) in outdoor cultures based on commercial size ponds. J World Aquac Soc. 2021 Oct 1;52(5):1047–58. [CrossRef]
- Fort A, Lebrault M, Allaire M, Esteves-Ferreira AA, McHale M, Lopez F, et al. Extensive Variations in Diurnal Growth Patterns and Metabolism Among Ulva spp. Strains. Plant Physiol . 2019 May 3 ;180(1):109–23. [CrossRef]
- Lapointe BE, Tenore KR. Experimental outdoor studies with Ulva fasciata Delile. I. Interaction of light and nitrogen on nutrient uptake, growth, and biochemical composition. J Exp Mar Biol Ecol. 1981 Jul 14;53[2–3]:135–52. [CrossRef]
- Ryther JH, Lapointe BE. The Effects of Nitrogen and Seawater Flow Rate on the Growth and Biochemical Composition of Gracilaria foliifera var. angustissima in Mass Outdoor Cultures. Botanica Marina. 1979 Jan 1 ;22(8):529–38. [CrossRef]
- Chopin T, Robinson SMC, Troell M, Neori A, Buschmann AH, Fang J. Multitrophic Integration for Sustainable Marine Aquaculture. In: Encyclopedia of Ecology, Five-Volume Set. Elsevier Inc.; 2008. p. 2463–75. [CrossRef]
- Neori A, Msuya FE, Shauli L, Schuenhoff A, Kopel F, Shpigel M. A novel three-stage seaweed (Ulva lactuca) biofilter design for integrated mariculture. J Appl Phycol. 2003 Nov ;15(6):543–53. [CrossRef]
- Buck BH, Shpigel M. ULVA: Tomorrow’s “Wheat of the sea”, a model for an innovative mariculture. J Appl Phycol . 2023 Jul 21 ;1:1–4. [CrossRef]
- Acién Fernández FG, Gómez-Serrano C, Fernández-Sevilla JM. Recovery of Nutrients From Wastewaters Using Microalgae. Front Sustain Food Syst. 2018 Sep 20; 2:59. [CrossRef]
- San Pedro A, González-López C V., Acién FG, Molina-Grima E. Outdoor pilot production of Nannochloropsis gaditana: Influence of culture parameters and lipid production rates in raceway ponds. Algal Res. 2015 Mar 1; 8:205–13. [CrossRef]
- Sharma AK, Sharma A, Singh Y, Chen WH. Production of a sustainable fuel from microalgae Chlorella minutissima grown in a 1500 L open raceway ponds. Biomass Bioenergy. 2021 Jun 1; 149:106073. [CrossRef]
- Shpigel M, Guttman L, Ben-Ezra D, Yu J, Chen S. Is Ulva sp. able to be an efficient biofilter for mariculture effluents? J Appl Phycol. 2019 Aug 1;31(4):2449–59. [CrossRef]
- Figueroa FL, Bonomi-Barufi J, Celis-Plá PSM, Nitschke U, Arenas F, Connan S, et al. Short-term effects of increased CO2, nitrate and temperature on photosynthetic activity in Ulva rigida (Chlorophyta) estimated by different pulse amplitude modulated fluorometers and oxygen evolution. J Exp Bot . 2021 Feb 2 ;72(2):491–509. [CrossRef]
- Figueroa FL, Korbee N, Abdala R, Jerez CG, López-de la Torre M, Güenaga L, et al. Biofiltration of fishpond effluents and accumulation of N-compounds (phycobiliproteins and mycosporine-like amino acids) versus C-compounds (polysaccharides) in Hydropuntia cornea (Rhodophyta). Mar Pollut Bull. 2012 Feb 1;64(2):310–8. [CrossRef]
- Aguilera J, De Gálvez MV, Conde R, Pérez-Rodríguez E, Viñegla B, Abdala R, et al. Series temporales de medida de radiación solar ultravioleta y fotosintética en Málaga. Actas Dermosifiliogr. 2004 Oct 1;95(1):25–31. [CrossRef]
- Green-Gavrielidis LA, Thornber CS. Will Climate Change Enhance Algal Blooms? The Individual and Interactive Effects of Temperature and Rain on the Macroalgae Ulva. Estuaries and Coasts . 2022 Sep 1 ;45(6):1688–700. [CrossRef]
- Xiao J, Zhang X, Gao C, Jiang M, Li R, Wang Z, et al. Effect of temperature, salinity and irradiance on growth and photosynthesis of Ulva prolifera. Acta Oceanologica Sinica. 2016 Oct 1;35(10):114–21. [CrossRef]
- Bews E, Booher L, Polizzi T, Long C, Kim JH, Edwards MS. Effects of salinity and nutrients on metabolism and growth of Ulva lactuca: Implications for bioremediation of coastal watersheds. Mar Pollut Bull. 2021 May 1; 166:112199. [CrossRef]
- Liu J, Tong Y, Xia J, Sun Y, Zhao X, Sun J, et al. Ulva macroalgae within local aquaculture ponds along the estuary of Dagu River, Jiaozhou Bay, Qingdao. Mar Pollut Bull. 2022 Jan 1; 174:113243. [CrossRef]
- Beer S, Eshel A. Photosynthesis of Ulva sp. II. Utilization of CO and HCO-3 when submerged. Vol. 70, Mar. Biol. Ecol. 1983. [CrossRef]
- Axelsson L, Ryberg H, Beer S. Two modes of bicarbonate utilization in the marine green macroalga Ulva lactuca. Plant Cell Environ . 1995 Apr 1 ;18(4):439–45. [CrossRef]
- Figueroa FL, Conde-Álvarez R, Gómez I. Relations between electron transport rates determined by pulse amplitude modulated chlorophyll fluorescence and oxygen evolution in macroalgae under different light conditions. Photosynth Res . 2003 ,75(3):259–75. [CrossRef]
- Figueroa FL, Israel A, Neori A, Martínez B, Malta E jan, Ang P, et al. Effects of nutrient supply on photosynthesis and pigmentation in Ulva lactuca (Chlorophyta): responses to short-term stress. Aquat Biol . 2009 Oct 22 ;7[1–2]:173–83. [CrossRef]
- Figueroa FL, Santos R, Conde-Álvarez R, Mata L, Gómez Pinchetti JL, Matos J, et al. The use of chlorophyll fluorescence for monitoring photosynthetic condition of two tank-cultivated red macroalgae using fishpond effluents. Botanica Marina . 2006 Sep 1 ;49(4):275–82. [CrossRef]
- Figueroa FL, Álvarez-Gómez F, Bonomi-Barufi J, Vega J, Massocato TF, Gómez-Pinchetti JL, et al. Interactive effects of solar radiation and inorganic nutrients on biofiltration, biomass production, photosynthetic activity and the accumulation of bioactive compounds in Gracilaria cornea (Rhodophyta). Algal Res. 2022 Nov 1; 68:102890. [CrossRef]
- Jerez CG, Malapascua JR, Sergejevová M, Masojídek J, Figueroa FL. Chlorella fusca (Chlorophyta) grown in thin-layer cascades: Estimation of biomass productivity by in-vivo chlorophyll a fluorescence monitoring. Algal Res. 2016 Jul 1; 17:21–30. [CrossRef]
- Figueroa FL, Conde-Álvarez R, Bonomi Barufi J, Celis-Plá PS, Flores P, Malta EJ, et al. Continuous monitoring of in vivo chlorophyll a fluorescence in ulva rigida (Chlorophyta) submitted to different CO2, nutrient and temperature regimes. Aquat Biol. 2014 Nov 20; 22:195–212. [CrossRef]
- Mata L, Schuenhoff A, Santos R. A direct comparison of the performance of the seaweed biofilters, Asparagopsis armata and Ulva rigida. J Appl Phycol . 2010 Feb 17;22(5):639–44. [CrossRef]
- Mata L, Santos R. Cultivation of Ulva rotundata (Ulvales, Chlorophyta) in raceways using semi-intensive fishpond effluents: yield and biofiltration. Proceedings of the 17th International Seaweed Symposium, Cape Town, South Africa, 28 January-2 February 2001. 2003;237–42.
- Msuya FE, Neori A. Effect of water aeration and nutrient load level on biomass yield, N uptake and protein content of the seaweed Ulva lactuca cultured in seawater tanks. J Appl Phycol . 2008 Dec 1 ;20(6):1021–31. [CrossRef]
- Msuya FE, Kyewalyanga MS, Salum D. The performance of the seaweed Ulva reticulata as a biofilter in a low-tech, low-cost, gravity generated water flow regime in Zanzibar, Tanzania. Aquaculture. 2006 Apr 28;254[1–4]:284–92. [CrossRef]
- Jiménez Del Río M, Ramazanov Z, García-Reina G. Ulva rigida (Ulvales, Chlorophyta) tank culture as biofilters for dissolved inorganic nitrogen from fishpond effluents. Hydrobiologia . 1996 ;326–327(1):61–6. [CrossRef]
- Neori A, Cohen I, Gordin H. Ulva lactuca Biofilters for Marine Fishpond Effluents II. Growth Rate, Yield and C:N Ratio. Botanica Marina . 1991 Jan 1;34(6):483–90. [CrossRef]
- Vandermeulen H, Gordin H. Ammonium uptake using Ulva (Chlorophyta) in intensive fishpond systems: mass culture and treatment of effluent. J Appl Phycol. 1990 Dec;2(4):363–74. [CrossRef]
- Debusk TA, Ryther JH, Hanisak MD, Williams LD. Effects of seasonality and plant density on the productivity of some freshwater macrophytes. Aquat Bot. 1981 Jan 1;10(C):133–42. [CrossRef]
- Bruhn A, Dahl J, Nielsen HB, Nikolaisen L, Rasmussen MB, Markager S, et al. Bioenergy potential of Ulva lactuca: Biomass yield, methane production and combustion. Bioresour Technol. 2011 Feb 1;102(3):2595–604. [CrossRef]
- Neori A, Shpigel M, Ben-Ezra D. A sustainable integrated system for culture of fish, seaweed and abalone . Vol. 186, Aquaculture. 2000. [CrossRef]
- Henley J. W, Levavasseur G, Franklin LA, Lindley ST, Ramus J, Osmond CB. Marine Ecology Progress Series. 1991. p. 75, 19–28 Diurnal responses of photosynthesis and fluorescence in Ulva rotundata acclimated to sun and shade in outdoor culture on JSTOR. Available from: https://www.jstor.org/stable/24825806.
- Henley WJ. Measurement and interpretation of photosynthetic light-response curves in algae in the context of photoinhibition and diel changes. J Phycol. 1993;29(6):729–39. [CrossRef]
- Wilhelm C, Becker A, Toepel J, Vieler A, Rautenberger R. Photophysiology and primary production of phytoplankton in freshwater. Physiol Plant. 2004 Mar 1;120(3):347–57. [CrossRef]
- Savvashe P, Mhatre-Naik A, Pillai G, Palkar J, Sathe M, Pandit R, et al. High yield cultivation of marine macroalga Ulva lactuca in a multi-tubular airlift photobioreactor: A scalable model for quality feedstock. J Clean Prod. 2021 Dec 20; 329:129746. [CrossRef]
- Zou D. The effects of severe carbon limitation on the green seaweed, Ulva conglobata (Chlorophyta). J Appl Phycol. 2014 Dec 1;26(6):2417–24. [CrossRef]
- Fort A, McHale M, Cascella K, Potin P, Usadel B, Guiry MD, et al. Foliose Ulva Species Show Considerable Inter-Specific Genetic Diversity, Low Intra-Specific Genetic Variation, and the Rare Occurrence of Inter-Specific Hybrids in the Wild. J Phycol. 2021 Feb 1;57(1):219–33. [CrossRef]
- Schneider G, Figueroa FL, Vega J, Chaves P, Álvarez-Gómez F, Korbee N, et al. Photoprotection properties of marine photosynthetic organisms grown in high ultraviolet exposure areas: Cosmeceutical applications. Algal Res. 2020 Aug 1; 49:101956. [CrossRef]
- Vega J, Bonomi-Barufi J, Gómez-Pinchetti JL, Figueroa FL. Cyanobacteria and Red Macroalgae as Potential Sources of Antioxidants and UV Radiation-Absorbing Compounds for Cosmeceutical Applications. Marine Drugs 2020, Vol 18, Page 659 . 2020 Dec 21 ;18(12):659. [CrossRef]
- Vega J, Álvarez-Gómez F, Güenaga L, Figueroa FL, Gómez-Pinchetti JL. Antioxidant activity of extracts from marine macroalgae, wild-collected and cultivated, in an integrated multi-trophic aquaculture system. Aquaculture. 2020 May 30; 522:735088. [CrossRef]
- R Ramírez T, Cortés D, Mercado JM, Vargas-Yañez M, Sebastián M, Liger E. Seasonal dynamics of inorganic nutrients and phytoplankton biomass in the NW Alboran Sea. Estuar Coast Shelf Sci. 2005 Dec 1;65(4):654–70. [CrossRef]
- Gomez Pinchetti JL, Del Campo Fernández E, Moreno Díez P, García Reina G. Nitrogen availability influences the biochemical composition and photosynthesis of tank-cultivated Ulva rigida (Chlorophyta). J Appl Phycol . 1998 ;10(4):383–9. [CrossRef]
- Figueroa FL, Bueno A, Korbee N, Santos R, Mata L, Schuenhoff A. Accumulation of Mycosporine-like Amino Acids in Asparagopsis armata Grown in Tanks with Fishpond Effluents of Gilthead Sea Bream, Sparus aurata. J World Aquac Soc . 2008 Oct 1 ;39(5):692–9. [CrossRef]
- Cabello-Pasini A, Aguirre-Von-Wobeser E, Figueroa FL. Photoinhibition of photosynthesis in Macrocystis pyrifera (Phaeophyceae), Chondrus crispus (Rhodophyceae) and Ulva lactuca (Chlorophyceae) in outdoor culture systems. J Photochem Photobiol B. 2000 Sep 1;57[2–3]:169–78. [CrossRef]
- Hanelt D, Figueroa FL. Physiological and Photomorphogenic Effects of Light on Marine Macrophytes. In 2012. p. 3–23. [CrossRef]
- Hendrickson L, Furbank RT, Chow WS. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence. Photosynth Res . 2004 ;82(1):73–81. [CrossRef]
- Figueroa FL, Domínguez-González B, Korbee N. Vulnerability and acclimation to increased UVB radiation in three intertidal macroalgae of different morpho-functional groups. Mar Environ Res. 2014 Jun 1; 97:30–8. [CrossRef]
- Figueroa FL, Bonomi Barufi J, Malta EJ, Conde-Álvarez R, Nitschke U, Arenas F, et al. Short-term effects of increasing CO2, nitrate and temperature on three mediterranean macroalgae: Biochemical composition. Aquat Biol. 2014 Nov 20; 22:177–93. [CrossRef]
- Longstaff BJ, Kildea T, Runcie JW, Cheshire A, Dennison WC, Hurd C, et al. An in situ study of photosynthetic oxygen exchange and electron transport rate in the marine macroalga Ulva lactuca (Chlorophyta). Photosynth Res. 2002 ;74(3):281–93. [CrossRef]
- Cabello-Pasini A, Figueroa FL. Effect of nitrate concentration on the relationship between photosynthetic oxygen evolution and electron transport rate in Ulva rigida (chlorophyta). J Phycol. 2005 Dec 16;41(6):1169–77. [CrossRef]
- Eilers PHC, Peeters JCH. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecol Modell. 1988 Sep 1;42[3–4]:199–215. [CrossRef]
- Morillas-España A, Lafarga T, Gómez-Serrano C, Acién-Fernández FG, González-López CV. Year-long production of Scenedesmus almeriensis in pilot-scale raceway and thin-layer cascade photobioreactors. Algal Res. 2020 Oct 1; 51:102069. [CrossRef]
- Shpigel M, Guttman L, Shauli L, Odintsov V, Ben-Ezra D, Harpaz S. Ulva lactuca from an Integrated Multi-Trophic Aquaculture (IMTA) biofilter system as a protein supplement in gilthead seabream (Sparus aurata) diet. Aquaculture. 2017 Dec 1; 481:112–8. [CrossRef]
- Castro R, Piazzon MC, Zarra I, Leiro J, Noya M, Lamas J. Stimulation of turbot phagocytes by Ulva rigida C. Agardh polysaccharides. Aquaculture. 2006 Apr 28;254[1–4]:9–20. [CrossRef]
- Valente LMP, Gouveia A, Rema P, Matos J, Gomes EF, Pinto IS. Evaluation of three seaweeds Gracilaria bursa-pastoris, Ulva rigida and Gracilaria cornea as dietary ingredients in European sea bass (Dicentrarchus labrax) juveniles. Aquaculture. 2006 Mar 1;252(1):85–91. [CrossRef]
- Abdala-Díaz RT, García-Márquez J, Rico RM, Gómez-Pinchetti JL, Mancera JM, Figueroa FL, et al. Effects of a short pulse administration of Ulva rigida on innate immune response and intestinal microbiota in Sparus aurata juveniles. Aquac Res. 2021 Jul 1;52(7):3038–51. [CrossRef]
- García-Márquez J, Rico RM, Sánchez-Saavedra M del P, Gómez-Pinchetti JL, Acién FG, Figueroa FL, et al. A short pulse of dietary algae boosts immune response and modulates fatty acid composition in juvenile Oreochromis niloticus. Aquac Res. 2020 Nov 1;51(11):4397–409. [CrossRef]
- Rico RM, Tejedor-Junco MT, Tapia-Paniagua ST, Alarcón FJ, Mancera JM, López-Figueroa F, et al. Influence of the dietary inclusion of Gracilaria cornea and Ulva rigida on the biodiversity of the intestinal microbiota of Sparus aurata juveniles. Aquaculture International. 2016 Aug 1;24(4):965–84. [CrossRef]
- García IB, Ledezma AKD, Montaño EM, Leyva JAS, Carrera E, Ruiz IO. Identification and Quantification of Plant Growth Regulators and Antioxidant Compounds in Aqueous Extracts of Padina durvillaei and Ulva lactuca. Agronomy 2020, Vol 10, Page 866 . 2020 Jun 18 ;10(6):866. [CrossRef]
- Hamouda RA, Hussein MH, El-Naggar NEA, Karim-Eldeen MA, Alamer KH, Saleh MA, et al. Promoting Effect of Soluble Polysaccharides Extracted from Ulva spp. on Zea mays L. Growth. Molecules 2022, Vol 27, Page 1394 . 2022 Feb 18 ;27(4):1394. [CrossRef]
- Sekhouna D, Kies F, Elegbede I, Matemilola S, Zorriehzahra J, Hussein EK. Potential assay of two green algae Ulva lactuca and Ulva intestinalis as bio-fertilizers. Sustainability, Agri, Food and Environmental Research . 2021 Mar 10 ;9(4). [CrossRef]
- Figueroa FL, Korbee N, Abdala-Díaz R, Álvarez-Gómez F, Gómez-Pinchetti JL, Acién FG. Growing algal biomass using wastes. Bioassays: Advanced Methods and Applications. 2018 Jan 1;99–117. [CrossRef]
- Neveux N, Bolton JJ, Bruhn A, Roberts DA, Ras M, Barre S La, et al. The Bioremediation Potential of Seaweeds: Recycling Nitrogen, Phosphorus, and Other Waste Products. 2018. [CrossRef]
- Nielsen MM, Bruhn A, Rasmussen MB, Olesen B, Larsen MM, Møller HB. Cultivation of Ulva lactuca with manure for simultaneous bioremediation and biomass production. J Appl Phycol. 2012 Jun;24(3):449–58. [CrossRef]
- Acién FG, Gómez-Serrano C, Morales-Amaral MM, Fernández-Sevilla JM, Molina-Grima E. Wastewater treatment using microalgae: how realistic a contribution might it be to significant urban wastewater treatment? Appl Microbiol Biotechnol . 2016 Nov 1 ;100(21):9013–22. [CrossRef]
- Posada JA, Brentner LB, Ramirez A, Patel MK. Conceptual design of sustainable integrated microalgae biorefineries: Parametric analysis of energy use, greenhouse gas emissions and techno-economics. Algal Res. 2016 Jul 1; 17:113–31. [CrossRef]
- Posadas E, Bochon S, Coca M, García-González MC, García-Encina PA, Muñoz R. Microalgae-based agro-industrial wastewater treatment: a preliminary screening of biodegradability. J Appl Phycol . 2014 Dec 1 ;26(6):2335–45. [CrossRef]
- Duarte CM, Wu J, Xiao X, Bruhn A, Krause-Jensen D. Can seaweed farming play a role in climate change mitigation and adaptation? Front Mar Sci. 2017 Apr 12;4(APR):245020. [CrossRef]
- Maulu S, Hasimuna OJ, Haambiya LH, Monde C, Musuka CG, Makorwa TH, et al. Climate Change Effects on Aquaculture Production: Sustainability Implications, Mitigation, and Adaptations. Front Sustain Food Syst. 2021 Mar 12; 5:609097. [CrossRef]
Figure 1.
pH vs. DO(%) (A), pH vs. Irradiance (B) and pH vs. Temperature (C) patterns at the raceway system during the experiments with different Ulva compressa culture densities (0.6, 0.8 and 1.0 kg FW m-2). Time 0 in X axis corresponds to 9.00 a.m. Open arrows correspond to midday; bold arrows indicates transitory pH increases in the afternoon.
Figure 1.
pH vs. DO(%) (A), pH vs. Irradiance (B) and pH vs. Temperature (C) patterns at the raceway system during the experiments with different Ulva compressa culture densities (0.6, 0.8 and 1.0 kg FW m-2). Time 0 in X axis corresponds to 9.00 a.m. Open arrows correspond to midday; bold arrows indicates transitory pH increases in the afternoon.
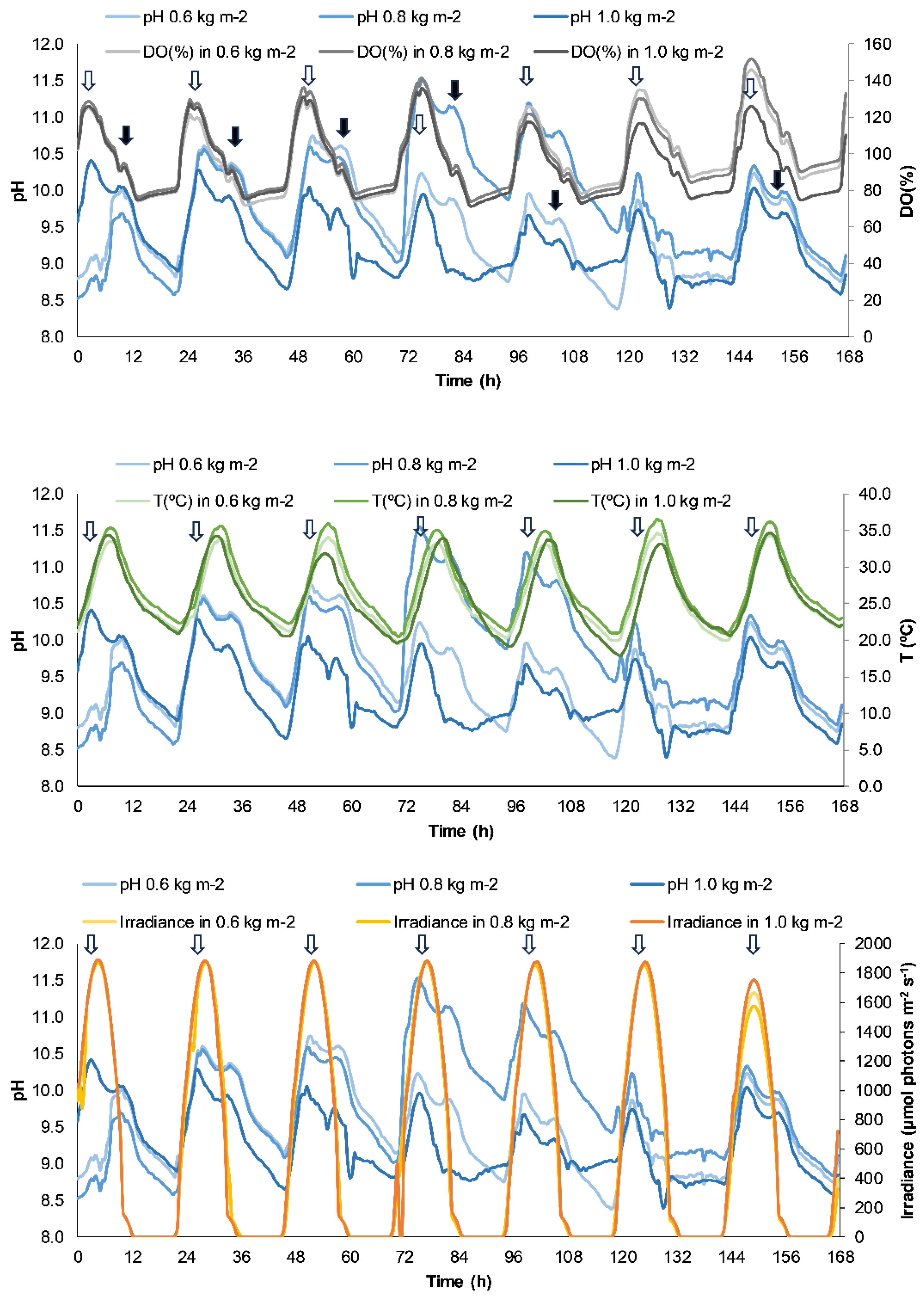
Figure 2.
Effective quantum yield (YII) (A) and in situ Electron Transport Rate (ETRin situ) (B) expressed in µmol electron m-2 s-1 measured in Ulva compressa growing in the raceway pond during the experiments at different culture density. Average irradiance level is also represented in secondary Y-axis. Different letters correspond to significant differences between treatments at a fixed time following one-way ANOVA (p < 0.05) after Tukey's test; lowercases corresponds to non-parametric Kruskal-Wallis test.
Figure 2.
Effective quantum yield (YII) (A) and in situ Electron Transport Rate (ETRin situ) (B) expressed in µmol electron m-2 s-1 measured in Ulva compressa growing in the raceway pond during the experiments at different culture density. Average irradiance level is also represented in secondary Y-axis. Different letters correspond to significant differences between treatments at a fixed time following one-way ANOVA (p < 0.05) after Tukey's test; lowercases corresponds to non-parametric Kruskal-Wallis test.
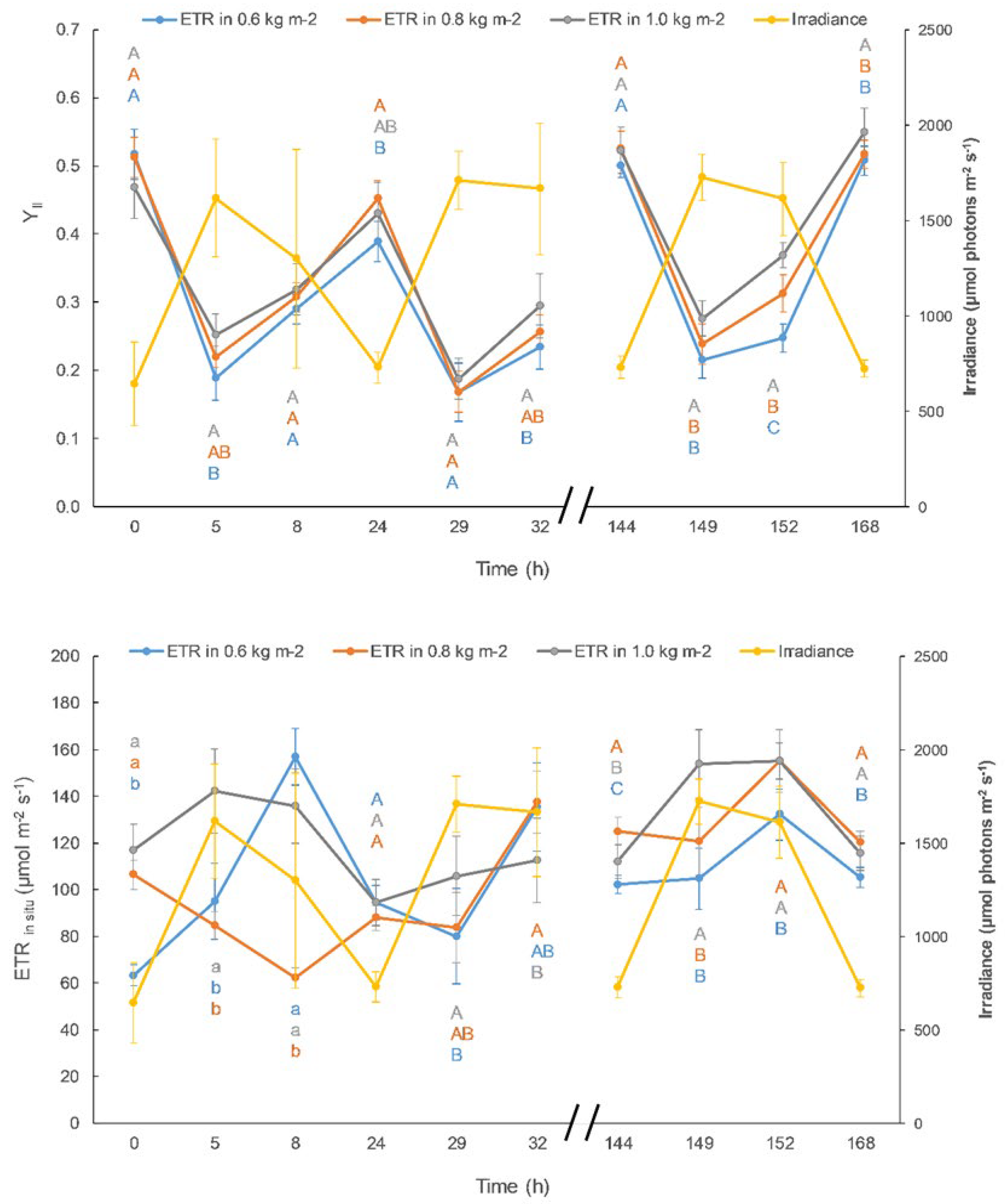
Figure 3.
YII (A) and ETRin situ expressed as µmol electron m-2 s-1 (B) in non-acclimatized and acclimatized Ulva compressa cultures at 1.0 kg FW m-2 density. Average irradiance level is also represented in secondary Y-axis. Different letters correspond to significant differences between treatments at a fixed time following t-student test (p < 0.05).
Figure 3.
YII (A) and ETRin situ expressed as µmol electron m-2 s-1 (B) in non-acclimatized and acclimatized Ulva compressa cultures at 1.0 kg FW m-2 density. Average irradiance level is also represented in secondary Y-axis. Different letters correspond to significant differences between treatments at a fixed time following t-student test (p < 0.05).

Figure 4.
Diagram showing the cultivation system used and the morphological changes observed in the algae. A: stocking tank; B: algae from the stocking tank; C: raceway reactor with the algae in culture; D: algae from the raceway reactor (note the change in size and colour).
Figure 4.
Diagram showing the cultivation system used and the morphological changes observed in the algae. A: stocking tank; B: algae from the stocking tank; C: raceway reactor with the algae in culture; D: algae from the raceway reactor (note the change in size and colour).
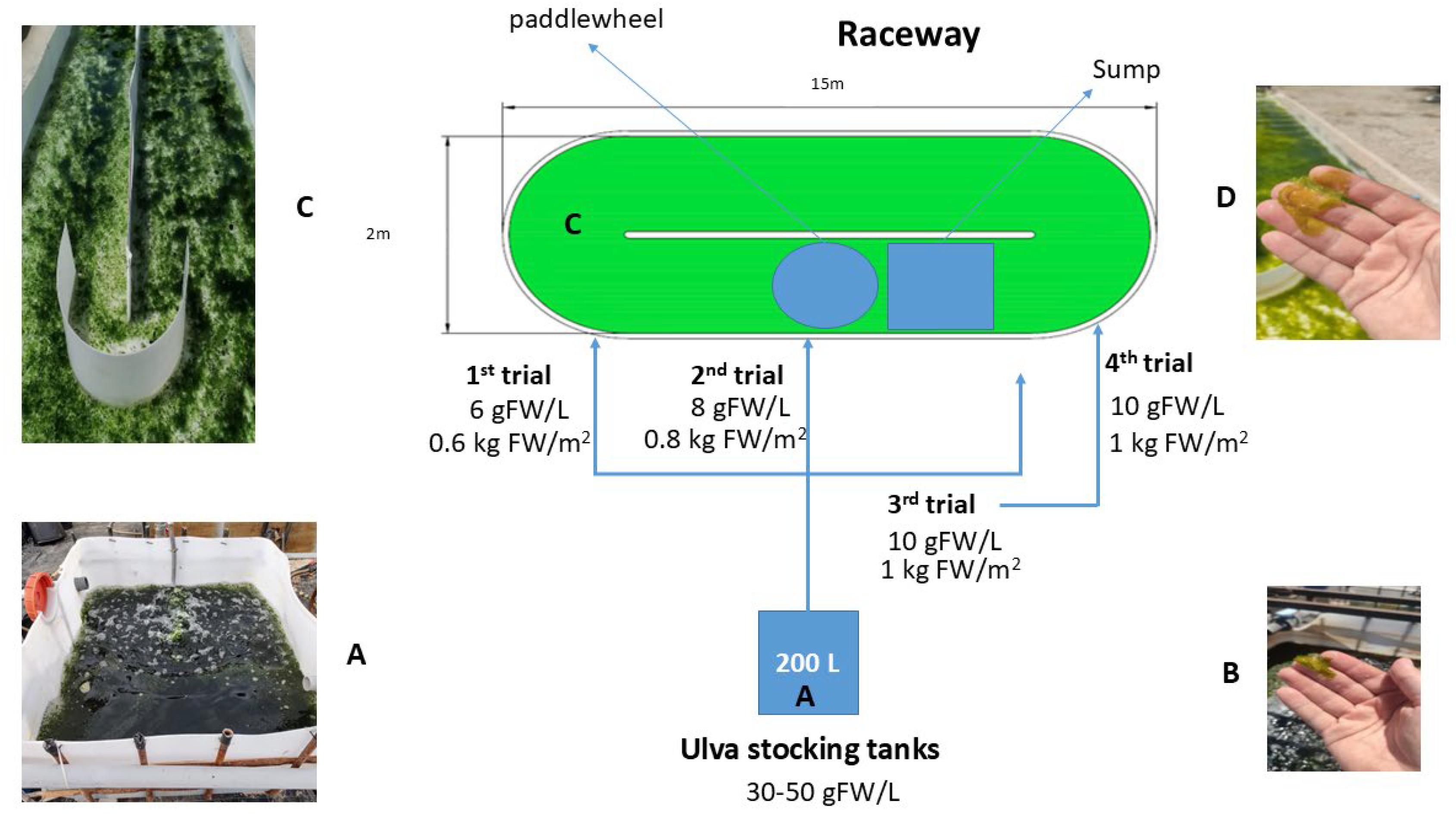

Table 1.
Growth parameters of Ulva compressa at different culture densities, determined at the end of every weekly experiment.
Table 1.
Growth parameters of Ulva compressa at different culture densities, determined at the end of every weekly experiment.
| Culture density | SGR (%day-1) |
Biomass production (kg FW) | Growth rate (g FW m-2 day-1) | Growth rate (g DW m-2 day-2) | Biomass production (kgFW/mg N) | Biomass production (kg FW/mg P) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.6 kg m-2 | 5.94 | 7.7 | 36.7 | 6.23 | 0.611 | 8.280 | ||||||
| 0.8 kg m-2 | 4.9 | 8.2 | 38.1 | 6.48 | 0.651 | 8.817 | ||||||
| 1.0 kg m-2 | 4.07 | 8.3 | 39.1 | 6.64 | 0.659 | 8.925 |
Table 2.
Photosynthetic parameters of Ulva compressa form Rapid Light Curves determined by using a Mini PAM fluorometer. Different letters correspond to significant differences between treatments following one-way ANOVA (p < 0.05) after Tukey's test.
Table 2.
Photosynthetic parameters of Ulva compressa form Rapid Light Curves determined by using a Mini PAM fluorometer. Different letters correspond to significant differences between treatments following one-way ANOVA (p < 0.05) after Tukey's test.
| Culture density | Fv/Fm | ETRmax (µmol m-2 s-1) | αETR (µmol electrons/µmol photons) | Ek (µmol photons m-2 s-1) | NPQmax | ETRmax/NPQmax | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.6 kg m-2 | 0.62 ± 0.010 B | 89.02 ± 12.16 A | 0.22 ± 0.008A | 412.00 ± 44.79 B | 0.74 ±0.074 C | 120.01 ± 15.34 A | ||||||
| 0.8 kg m-2 | 0.63 ± 0.010 B | 90.51 ± 11.62 A | 0.22 ± 0.034A | 352.81 ± 15.23 B | 1.41 ± 0.044 B | 64.31 ± 27.03 B | ||||||
| 1.0 kg m-2 | 0.68 ± 0.007 A | 94.82 ±5.69 A | 0.18 ± 0.009A | 512.46 ± 36.22 A | 1.51 ± 0.028 A | 62.69 ± 4.79 B |
Table 3.
Growth parameters of Ulva compressa acclimatized during 1 week at the photobioreactor conditions, determined at the end of the experiment.
Table 3.
Growth parameters of Ulva compressa acclimatized during 1 week at the photobioreactor conditions, determined at the end of the experiment.
| Culture density | SGR(%day-1) | Biomass production (kg FW) | Growth rate (g FW m-2 day-1) | Growth rate (g DW m-2 day-1) | Biomass production (kg FW/mg N) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pre-acclimatized | 4.07 | 8.30 | 38.10 | 6.48 | 0.66 | |||||
| Post-acclimatized | 4.80 | 10.1 | 47.62 | 8.10 | 0.79 |
Table 4.
Photosynthetic parameters of U. compressa derived from RLCs performed on acclimated algae and non-acclimated algae at 1.0 kg FW m-2 density. Different letters correspond to significant differences between treatments following t-student test (p < 0.05).
Table 4.
Photosynthetic parameters of U. compressa derived from RLCs performed on acclimated algae and non-acclimated algae at 1.0 kg FW m-2 density. Different letters correspond to significant differences between treatments following t-student test (p < 0.05).
| Algae condition |
Fv/Fm | ETRmax (µmol m-2 s-1) |
αETR (µmol electrons /µmol photons) | Ek (µmol photons m-2 s-1) | NPQmax | ETRmax/NPQmax (µmol m-2 s-1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-acclimation | 0.680 ± 0.007B | 94.89 ± 5.69B | 0.180 ± 0.009B | 512.460 ± 36.221B | 1.510 ± 0.028A | 62.69 ± 4.79B | ||||||
| Post-acclimation | 0.740 ± 0.006A | 184.73 ± 28.52A | 0.230 ± 0.009A | 852.340 ± 99.315A | 0.750 ± 0.027B | 245.98 ± 45.85A |
Table 5.
Cultivation conditions, biomass yields, biofiltering efficiency and nitrogen removal of Ulva spp. cultivated in different experimental systems. Modified from [26].
Table 5.
Cultivation conditions, biomass yields, biofiltering efficiency and nitrogen removal of Ulva spp. cultivated in different experimental systems. Modified from [26].
| Specie | Tank Volume (L) | Stocking density (kg FW m-2) | Growth (g L-1 day-1) | Water exchange (L day-1) | Growth rate (g DW m-2 day-1) | References | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U. compressa | 3000 | 0,6-1 | 0.37-0.48 | 0 | 6,23-8 | This study | ||||||
| U. pseudorotundata | 200 | 1.2 | Not | 0 | 7.5-8 | [6] | ||||||
| U. lactuca | 800 | 1-3 | 0.32-0.17 | 0 | 25-13 | [26] | ||||||
| U. rigida | 110 | 1.9 | Not | 2,4-96 | 44-73 | [42] | ||||||
| U. rigida | 1900 | 1.9 | Not | 14,4 | 48 | [43] | ||||||
| U. lactuca | 600 | 1 | 0.19-0.63 | 34 | 11-38 | [44] | ||||||
| U. reticulata | 40 | 1 | 1.35-2.3 | 2040 | 46 | [45] | ||||||
| U. rigida | 750 | 2.5 | 0.09-0.32 | 2-12 | 40 | [46] | ||||||
| U. lactuca | 600 | 2-6 | 0.24-0.42 | 4-16 | 55 | [47] | ||||||
| U. lactuca | 600 | 1 | Not | 4-8 | 55 | [48] | ||||||
| U. lactuca | 1700 | 1 | Not | 1-24 | 45-16 | [49] | ||||||
| U. lactuca | 600 | 1-8 | 0.37-0.16 | 12 | 12,32 | [50] | ||||||
| U. lactuca | 600 | 1.5 | 0.39 | 2 | 21,3 | [51] | ||||||
| U. lactuca | 900-1700 | 1 | 0.26-0.64 | 14-56 | 19 | [21] |
Table 6.
Values of Pearson correlations values among the following parameters: SGR (%), ETRmax, ETRmax/NPQ and Nitrate Uptake Rate (NUR). Bold letters indicate positive significant values (* p < 0.05; ** p > 0.01).
Table 6.
Values of Pearson correlations values among the following parameters: SGR (%), ETRmax, ETRmax/NPQ and Nitrate Uptake Rate (NUR). Bold letters indicate positive significant values (* p < 0.05; ** p > 0.01).
| SGR (%) | ETRmax (µmol m-2 s-1) | ETRmax/NPQmax (µmol m-2 s-1) | NUR (µmol N g-1 DW h-1) | |||||
|---|---|---|---|---|---|---|---|---|
| SGR (%) | - | -0.2689 | 0.8147** | 0.9969** | ||||
| ETRmax | - | - | 0.0772 | -0.2605 | ||||
| ETRmax/NPQmax | - | - | - | 0.8353** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



