Submitted:
09 December 2025
Posted:
11 December 2025
You are already at the latest version
Abstract
Dengue outbreaks highlight the need to quantify how environments sustain populations of the mosquito vectors Ae. aegypti and Ae. albopictus. Most control activities focus on households, while non-residential premises receive little attention. We conducted a cross-sectional ovitraps survey in August 2025 in Tapachula, Chiapas, Mexico, deploying 593 ovitraps: 297 in residential neighborhoods and 293 in non-residential sites. We collected 42,758 eggs; 61.5% (n = 26,316) came from residential areas and 38.5% (n = 16,442) from non-residential areas. Ovitrap positivity was higher in residential versus in non-residential areas (92.6% vs. 87.0%; p = 0.025), and the mean number of eggs per ovitrap (88.6 ± 5.1 vs. 56.1 ± 3.3 eggs; p < 0.001), yielding 298.0 vs. 175.3 eggs/ha, respectively. From a subsample of 16,965 eggs, 7,543 adults emerged (44.5%), dominated by Ae. aegypti in residential areas (83.3%) and Ae. albopictus in non-residential areas (80.0%). Vector burden differed by species: Ae. aegypti densities were higher in residential areas (51.0 vs. 4.5 adults/ha), whereas Ae. albopictus densities were higher in non-residential sites (18.2 vs. 10.2 adults/ha). These patterns show that both residential and non-residential premises are important sources of dengue vectors and should be included in integrated surveillance and control strategies.
Keywords:
Aedes aegypti
; Ae. albopictus
; dengue
; entomological surveillance
; oviposition
; ovitraps
; vector control
1. Introduction
In 2024, Latin America experienced the largest dengue outbreak on record, with 13,063,434 suspected cases, while Mexico reported 558,846 cases of which 125,160 were confirmed [1,2]. The factors attributed to these alarming results include a lack of effective vaccines, limited development of antiviral drugs, and climate change [3]. However, we must also recognize that field methods for controlling the Ae. aegypti and Ae. albopictus vectors typically result only in temporary reductions in their populations [4]. This is likely due to insecticide resistance in dengue vectors, Ae. aegypti and Ae. albopictus [5,6,7]. In addition, many non-residential areas, such as parks, cemeteries, and schools, are never treated with insecticides. These areas are vast and complex to access. These residual populations in non-residential areas often re-infest the same areas and the residential area in subsequent seasons. Few studies have quantified the epidemiological impact of domestic or household vector populations, and even fewer have examined populations outside of human dwellings [8]. A likely reason why these vectors persist in non-residential urban areas is that, in the past, entomological indices, such as the House Index used during yellow fever campaigns, focused exclusively on mosquito populations inside homes [9]. These Aedine indices, along with Container and Breteau indices, were incorporated as dengue epidemics emerged at the end of the 1970s. Today, many programs still believe that dengue transmission only occurs within households, leading most control efforts to focus there. In recent years, reports on outdoor cases and vectors have increased. For example, [10] collected Ae. aegypti in school classrooms infected with Dengue serotype 2 in Mérida, Yucatán, Mexico. [11] documented a high prevalence and productivity of Ae. aegypti pupal populations in non-residential sites in Iquitos, Peru, such as markets and riverboats, with values comparable to or higher than those in residential areas (≈122–213 vs. 122–161 pupae/ha). In Monterrey, Mexico, Zarate-Nahon et al. (2013) found that 94. 3% of Ae. aegypti were present at non-residential sites such as schools, commercial areas, churches, tire repair shops, factories, bus stations, meat processing facilities, libraries, and recreational sites. Finally, [12] found that non-residential environments were more likely to be productive sites for Ae. aegypti than residential sites. Vacant lots are often located near homes, have abundant vegetation, and contain discarded items that retain water and can serve as breeding grounds for Ae. aegypti larvae.
Considering the potential significance of uncontrolled vector populations outside homes, this study aimed to: 1) compare the egg productivity of Aedes spp. between residential and non-residential areas; 2) identify and compare the species composition of Ae. aegypti and Ae. albopictus in residential and non-residential neighborhoods; 3) Compare the vector burden of Aedes species between residential and non-residential sites.
2. Materials and Methods
2.1. Study Area
This study was conducted in Tapachula, Chiapas, a city located in southeastern Mexico with 353,706 inhabitants (2020 census) (Figure 1). Between the parallels 14°37’ and 15°15’ N, the meridians 92°09’ and 92°28’ W, altitude between 0 and 2,700 m, and it borders Guatemala to the south. The region has a humid tropical climate, with an average annual temperature of 28°C and a relative humidity of 80%. Annual rainfall is characterized by a rainy season from May to October and a dry season from November to April, with annual precipitation ranging between 1,200 – 5,000 mm [13]. In addition to its geographic location, which favors the reproduction and spread of the Ae. aegypti mosquito, resulting in annual outbreaks of dengue fever, this region has been characterized by intense migratory movements from Central and South America to the United States [14].
2.2. Egg Collection
Mosquito eggs were collected in both residential and non-residential areas during August 2025. The sampling period encompassed three weeks and was conducted in the rainy season. The sites chosen for ovitrap placement were divided into two categories: residential, such as Neighborhood Laureles, Neighborhood 5 de Febrero, and Neighborhood Raymundo Enríquez, which are primarily inhabited areas; and non-residential, including sites as: Cementeries, public parks, schools, vacant lots, and other sites (car junk lot, mechanical workshop, car wash, and tire repair shop) [11,15]. Four pairs of field technicians were assigned to each study site to place ovitraps. A map divided each site into four sections, with each team responsible for one section. When ovitrap placement was permitted, each trap was georeferenced using the Google Earth® app on the field technicians' cell phones. In the laboratory, all georeferences for each ovitrap were collected and their coordinates verified on a single Google Earth map. Finally, the coordinates were converted into KMZ format on the MyGeodata platform [16] (https://mygeodata.cloud/converter/kmz-to-kml). A total of 300 ovitraps were distributed across the residential areas. Based on accessibility, 23, 25, and 31 city blocks were selected from the three neighborhoods: 5 de Febrero, Raymundo Enríquez, and Laureles, respectively. Within each block, four ovitraps were placed, one on each block side. The ovitraps were distributed within households in the living room, dining room, front yard, and backyard. Similarly, 300 ovitraps were deployed in non-residential areas. They were positioned in safe locations to prevent disturbance or damage by people or animals. The ovitraps used were the standard design; i.e., black plastic containers, 7 cm in diameter and 16 cm in height, with a capacity of 500 ml [17]. A strip of white filter paper (7 cm wide x 15 cm long) was used as an oviposition substrate to collect eggs of dengue vectors. This strip was lined on the inner container walls. Ovitraps were filled with 400 ml of water, enough to keep the filter paper moist for at least 10 days and prevent evaporation caused by the high temperatures in the study area. The filter paper from each ovitrap was removed weekly from the date of placement and transferred to the CRISP/INSP insectary in plastic bags to determine egg positivity, egg count, and egg hatching for larval development and to identify the species present in the adult stage.
2.3. Egg Counts and Aedes Identification
Individual strips of filter paper from ovitraps were placed under a dissecting microscope for egg counting. Subsequently, strips were transferred to 1-L containers of dechlorinated water to stimulate egg hatching. After 12 -24 h, the first-instar larvae were fed by pouring fine-ground mouse food (Lab Diet®). The photoperiod in the insectary was 12:12 Light: Dark, with an average temperature of 25 ± 1 °C and a relative humidity of 40 ± 10% [18]. Emergent adults were identified based on morphological characteristics described by [19].
2.4. Data Analysis
We addressed three main objectives. First, to compare egg productivity of Aedes sp. between residential and non-residential areas, we calculated the proportion of ovitraps with eggs (ovitrap positivity) and the mean number of eggs per ovitrap for each area. Differences in ovitrap positivity between residential and non-residential areas were evaluated using the chi-square test of independence. Because egg counts per ovitrap were highly skewed and did not meet normality assumptions, we compared egg productivity between residential and non-residential areas using the Mann–Whitney U test. Within residential areas, differences in egg counts among neighborhoods (5 de Febrero, Raymundo Enríquez, and Laureles) were assessed using the Kruskal–Wallis test followed by Dunn’s post hoc test with p-value adjustment for multiple comparisons. Within non-residential areas, differences in egg productivity among site categories (schools, vacant lots, public parks and other sites) were evaluated using the Kruskal–Wallis test followed by the Dwass–Steel–Critchlow–Fligner (DSCF) post hoc test. To complement these non-parametric analyses, we additionally provide boxplots showing medians and interquartile ranges in supplementary Figures S1–S3 [20].
Second, to identify and compare the species composition of Ae. aegypti and Ae. albopictus between residential and non-residential areas, we constructed contingency tables of species by area type (residential vs. non-residential), by residential neighborhood, and by non-residential site category. Associations between species and each spatial factor were evaluated using chi-square tests of independence. Standardized Pearson residuals were examined to identify species that were over- or under-represented in each neighborhood or non-residential site category.
Third, to describe and compare the vector burden of Aedes species between residential and non-residential sites, we defined vector burden as the density of eggs and emerged adults per hectare. For each area (residential and non-residential), we calculated total egg counts and total numbers of emerged Ae. aegypti and Ae. albopictus adults and expressed these values as eggs per hectare and adults per hectare. These density measures were compared descriptively between residential and non-residential areas, because they represent area-level indices rather than independent sampling units suitable for formal hypothesis testing. All statistical analyses were performed using Jamovi version 2.6.44 and R version 4.5.2.
3. Results
Of the 600 ovitraps initially placed, seven could not be located, so 593 were examined. Of these, 297 (50.1%) were installed in households within residential areas, while the remaining 293 (49.9%) were placed in various non-residential locations. These included four cemeteries (119 traps, 20%), three public parks (45 traps, 7.6%), nine schools (39 traps, 6.6%), and 26 vacant lots (26 traps, 4.4%). The other places considered as “other sites” include: two car junk lots (31 traps, 5.2%), 29 mechanical workshops (29 traps, 4.9%), 26 vacant lots (26 traps, 4.4%), two car washes (5 traps, 0.8%), and one tire repair shop (2 traps, 0.3%) (Table 1).
In residential areas, 92.6% (275/297) of ovitraps tested positive, compared to 87.0% (255/293) in non-residential areas. The higher positivity rate in residential zones was statistically significant, with a difference of 5.6 percentage points (χ² = 4.99, df = 1, p = 0.025).
A total of 42,758 eggs were collected across all study areas, 61.5% (n = 26,316) from residential zones and 38.5% (n = 16,442) from non-residential zones. Ovitraps in residential areas yielded a mean of 88.6 (SE ±5.1) eggs, significantly larger than non-residential areas with 56.1 (SE ±3.3) eggs (U = 33417; p < 0.001) (Figure 2 and Figure S1).
Regarding the residential area, the mean number of eggs per ovitrap was 73.9 (SE ± 9.17) in 5 de Febrero; 114.3 (SE ± 9.7) in Raymundo Enríquez, and 78.0 (SE ± 6.87) in Laureles. The nonparametric Kruskal-Wallis test indicated significant differences in the average number of eggs across the three neighborhoods (H = 16.084; p < 0.001). Dunn's post hoc test demonstrated that Raymundo Enríquez had a significantly higher mean number of eggs than 5 de Febrero (p = 0.0003) and Laureles (p = 0.018). In contrast, no significant differences were observed between 5 de Febrero and Laureles (p = 0.729) (Figure 3 and Figure S2).
In non-residential areas, the mean number of eggs collected varied across site types. Schools had the highest means, with 74.4 (SE ± 9.2) eggs, indicating more favorable conditions for mosquito oviposition in this environment. Cemeteries had a mean of 56.6 (SE ± 5.4) eggs per ovitrap, while public parks slightly outperformed, averaging 58.5 (SE ± 7.4) eggs per ovitrap. Car junk lots, Mechanical workshops, car washes, tire repair shops, grouped under the "other" category, showed the lowest egg mean of 38.1 (SE ± 4.9). The non-parametric Kruskal-Wallis test demonstrated significant differences in the quantity of eggs among site categories (χ² = 14.1; df = 4; p = 0.007). Post-hoc analysis using the Dwass-Steel-Critchlow-Fligner multiple comparisons test revealed a significant difference between schools and the “other” category (W = –5.019; p = 0.004), with no significant differences in the other pairwise comparisons (p > 0.05). In summary, these findings indicate that mosquito egg productivity is higher in schools compared to sites classified as “other” (Figure 4 and Figure S3).
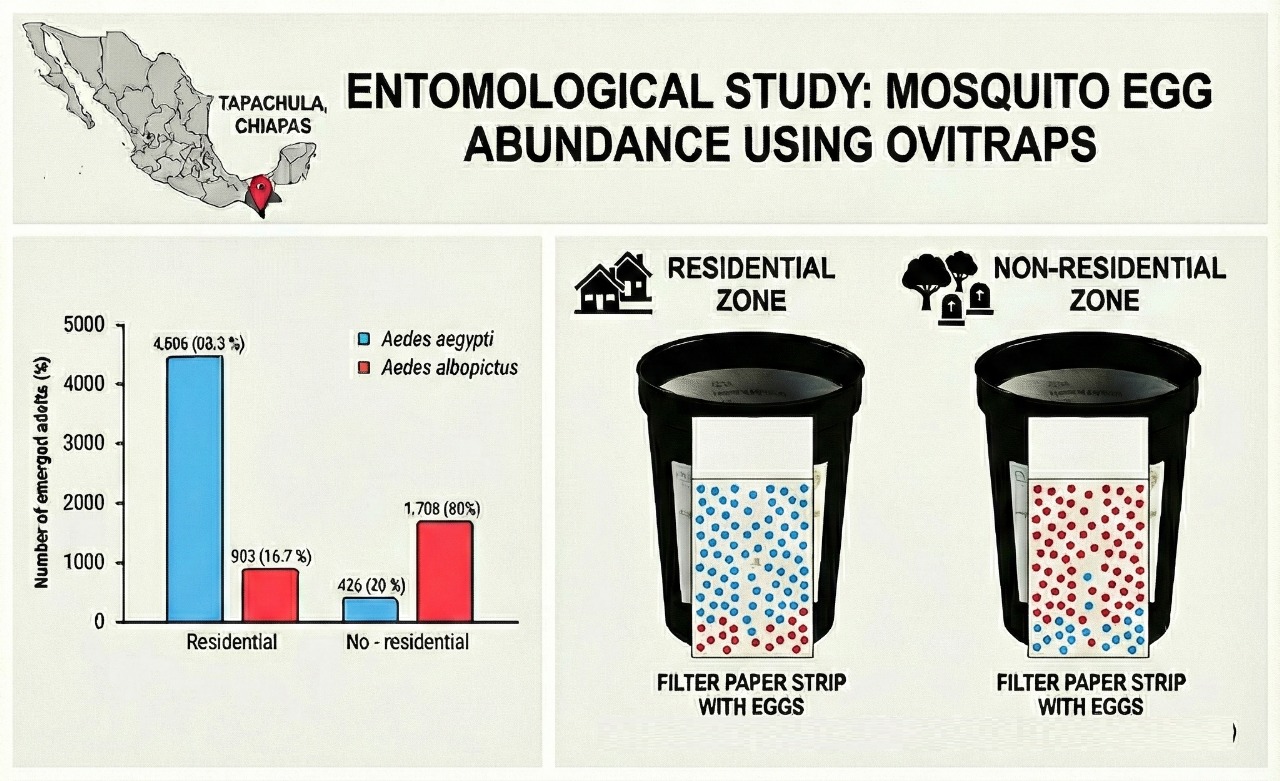
Out of a total of 42,758 eggs collected, a subset of 16,965 eggs was selected for species identification purposes. Of these, 7,543 (44.5 %) adults emerged and were subsequently identified. Among them Ae. aegypti constituted 65.4% (n = 4,932), whereas Ae. albopictus accounted for 34.6% (n = 2,611). Regarding their origin, 5,409 adults (71.7%) were from residential areas, while 2,134 (28.3%) were from non-residential areas. In residential zones, Ae. aegypti exhibited a predominant presence at 83.3% (n=4,506), in contrast to Ae. albopictus, which represented 16.7% (n = 903). Conversely, in non-residential zones, the species composition was inverted: Ae. aegypti comprised only 20.0% (n = 426), whereas Ae. albopictus constituted 80.0% (n=1,708). The chi-square statistical test (χ² = 2,734; df = 1; p < 0.001) demonstrated a highly significant association (Figure 5).
3.1. Frequency of Emerged Species in Residential Sites
In total, 5,409 adults emerged from the eggs collected in residential areas, of which 83.3% belonged to Ae. aegypti (n = 4,506) and 16.7% to Ae. albopictus (n = 903). The productivity of both species varied significantly across neighborhoods (Figure 5). The chi-square test of independence revealed a highly significant association between species and neighborhood (χ² = 796; df = 2; p < 0.001).
Across the residential areas, Ae. aegypti was the predominant species compared to Ae. albopictus. Pearson's standardized residuals showed that Ae. aegypti was overrepresented in the 5 de Febrero (residual = 4.86) and Laureles (residual = 4.42) neighborhoods, whereas Ae. albopictus was strongly underrepresented in both areas (residuals = -10.85 and -9.86, respectively). Although in absolute numbers Ae. aegypti was more abundant in the three neighborhoods (Figure 6), and the standardized residues show proportional differences. In Raymundo Enríquez, Ae. albopictus represented a much larger proportion than expected, which explains its overrepresentation (residual = 21.17), while Ae. aegypti was notably underrepresented (residual = -9.48) even though the absolute number of Ae. aegypti is still higher.
3.2. Frequency of Emerged Species in Non-Residential Areas
From the eggs collected at non-residential sites, a total of 2,134 adult mosquitoes emerged. Of these, 80.0% were identified as Ae. albopictus (n = 1,708), and 20.0% as Ae. aegypti (n = 426). The number of emerging adults was not evenly distributed among the various types of urban sites (Figure 7). In all sites, Ae. albopictus was the predominant species. The most abundant sites for Ae. albopictus were cemeteries and public parks, with 81.8% and 72.4%, respectively. Similarly, Ae. aegypti showed high adult abundance in cemeteries and public parks with 18.2% and 27.6%, respectively. A chi-square test revealed significant differences in species composition based on site type (χ² = 19.6; df = 3; p < 0.001). Post-hoc analysis based on standardized residues showed that, among non-residential sites, public parks are particularly favorable environments for producing Ae. aegypti, while Ae. albopictus is generally dominant in most other non-residential sites evaluated.
3.3. Vector Burden by Residential and Non-Residential Areas
Our research encompassed 88.3 hectares in residential zones and 93.8 hectares in non-residential zones. The residential zones yielded a total of 26,316 eggs, averaging 298.0 eggs per hectare. Conversely, the non-residential zones produced 16,442 eggs, averaging 175.3 per hectare. Species productivity varied between site types and species. For Ae. aegypti, the adult density per unit area was much higher in the residential zone (51.0 adults/ha) than in non-residential sites (4.5 adults/ha). Conversely, for Ae. albopictus, non-residential sites had a productivity of 18.2 adults/ha, while residential areas reached 10.2 adults/ha.
4. Discussion
Despite unsubstantiated suspicions attributing inadequate control rates in both indoor and outdoor house environments against Ae. aegypti, the factors explaining these findings remain poorly documented. The recurring epidemics in Latin America annually necessitate consideration of the possibility of additional vector populations existing outside human dwellings, thereby eluding vector control measures. This study, conducted in southern Mexico, generated data that support these suspicions.
One of the most efficient methods for monitoring the presence, abundance, and composition of Aedes sp. in specific times and spaces is the use of ovitraps. The ovitraps can be placed inside or around homes in residential areas—and in non-residential areas such as cemeteries, vacant lots, or recreational parks [21]. Information obtained from ovitraps helps determine the risk of transmission of dengue, Zika, or Chikungunya viruses. In the present study, the high ovitrap positivity for Aedes sp. eggs indicates the presence of gravid females feeding on a blood source near or inside homes. Our results showed that Ae. aegypti was significantly predominant compared to Ae. albopictus in the three selected residential areas. Furthermore, the pattern of species presence changed in non-residential areas, where Ae. albopictus showed a significantly greater presence than Ae. aegypti. This pattern was different in a study of entomological surveillance conducted in Brazil, where the densities of Ae. aegypti were higher than those of Ae. albopictus in residential and non-residential areas [22].
In our study, egg productivity was significantly higher in residential than in non-residential sites (88.6 ± 5.1 vs. 56.1 ± 3.3 eggs/ovitrap, p < 0.0001; Figure 2). This finding indicates that, on average, dwellings harbor the highest density of Aedes spp. eggs, and consequently, the vector burden was more significant in residential sites, while non-residential sites contribute a smaller, but still important, portion of total egg production. Similarly, [22] reported in Rio de Janeiro that, using ovitraps in three neighborhoods with different socio-environmental characteristics, an average of 69 eggs per dwelling per week, compared to 44 eggs per non-residential property per week. An increasing number of reports documenting higher activity of dengue vectors in non-residential urban environments highlight these areas as supplementary targets for control. In Yucatán, Mexico, 12 assessed the role of vacant lots as sources of immature Ae. aegypti: 74% of immatures were found in non-residential premises, and vacant lots represented 50% of the non-residential area, whereas 26% of immature stages were located in residential premises. In a subsequent study, [8] found freshly blood-fed Ae. aegypti females in Catholic church buildings, with 97.4% of blood meals identified as human.
Regarding the impact of both Aedes species on vector burden across residential and non-residential premises in Tapachula, Chiapas, Mexico, our data revealed clear patterns. Populations of Ae. aegypti were consistently dominant across all residential sites, whereas Ae. albopictus predominated in all non-residential sites (Figure 5). In contrast, [22] reported that Ae. aegypti remained the predominant species in both residential and non-residential premises, with Ae. albopictus represents only a minor fraction of adults, even in non-residential settings. Thus, while in Rio de Janeiro, non-residential premises mainly reinforced an Ae. aegypti driven vector burden, in Tapachula non-residential environments, appears to contribute to the production of Ae. albopictus increasing the heterogeneity of the local vector community.
This situation occurring in Southern Mexico represents an expansion trend of Ae. aegypti from household indoor spaces to nearby and surrounding inhabited spaces, and Ae. albopictus has a distribution frequency toward outdoor, densely wooded surroundings. This is explained by the sequential phases of adaptation of invasive species during colonization of a new geographical territory. Nevertheless, the epidemiological importance of both vector species is well known elsewhere, along with their role in driving larger and more frequent dengue and other arboviral outbreaks in Mexico and other endemic countries. Indeed, vector control programs should consider integrating non-residential premises to achieve more substantial reductions in vector populations. Even more, there is an urgent need to develop specific research targeting field operations and strategies. For instance, traditional cleaning campaigns are popular and recommended to eliminate breeding sites in backyards. Even though removal of small to medium-sized containers is successful, this operation is not applied to cemeteries, public parks, schools, vacant lots, car junk lots, markets, churches, body shops, etc. In non-residential areas, the civilian population does not conduct any control measures; therefore, free mosquito certification needs to be strongly regulated by the City Health Departments. In addition, the importance of non-residential areas is increased by the report of dormant eggs [23] attached to objects within them, such as the inner walls of discarded tires and other commonly used containers like plastic buckets. Similarly, dry eggs are an important support for the vertical transmission of dengue viruses during the interepidemic season. [24] reported that around 5% of adults emerged from field-collected Ae. aegypti eggs were positive for dengue viruses from Chiapas, Oaxaca, Guerrero, and Veracruz.
This study showed the vector burden of Ae. aegypti and Ae. albopictus in residential and non-residential areas. This will be useful for making substantial adjustments to modern integrated vector control strategies, as sustainable, area-wide field operations are urgently demanded.
5. Conclusions
This study shows that urban residential areas in Tapachula, Chiapas, sustain a higher overall egg productivity (mean of 88.6 eggs) of Aedes spp. than non-residential premises (mean of 56.1 eggs), as reflected by the greater ovitrap positivity and mean number of eggs per trap in households (Figure 2). In contrast, non-residential sites such as schools, vacant lots, public parks, and cemeteries accounted for a substantial fraction (38.5%) of total egg production and were particularly important for Ae. albopictus (Figure 5). Species composition differed between environments: Ae. aegypti predominated in all residential areas (Figure 6), whereas Ae. albopictus was consistently more abundant in non-residential sites, resulting in distinct area-level patterns of vector burden per hectare for each species. These findings indicate that dengue vector control programs cannot rely solely on household-based interventions. Non-residential premises in the urban landscape act as sources of vectors that can re-infest nearby homes and public spaces. Future studies of the role of Aedes sp. populations outside households are recommended to incorporate an area-wide strategy into current integrated dengue vector management programs.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Comparison of total egg counts between residential and non-residential areas, Figure S2: Distribution of total egg counts across Residential sites, Figure S3: Distribution of total egg counts across Non-residential sites.
Author Contributions
Conceptualization, I.F.-S, C.V.-T., J.G.O.-G., and K.M.V.-D.; methodology, J.G.O.-G., C.V.-T., K.M.V.-D., F.Z.V.-M., J.C.R.-D. and R.M.S.-C.; software, L.A.C.-V. and D.A.M.-L.; formal analysis, L.A.C.-V. and D.A.M.-L.; investigation, C.V.-T., I.F.-S. J.G.O.-G., K.M.V.-D., and F.Z.V-M.; writing—review and editing, all authors; visualization, F.Z.V.-M.; supervision, C.V.-T., I.F.-S., J.G.O.-G., K.M.V.-D., and F.Z.V-M.; all authors have read and agreed to the published version of the manuscript.
Funding
This work was made possible by the financial support of institutional resources from the National Institute of Public Health (INSP).
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
To QFB Luis Alonso Escobar Pérez, Victor Hugo López, and Virginia López González for supporting the mass breeding of Ae. aegypti under insectary conditions. To the technicians members of the CRISP Insecticide Evaluation Unit, René Monzón Vera, Eleazar Pérez Gómez, Carlos Chamlati Morán, Odette De Los Santos Domínguez, José Alfredo Rodas Mazariegos, Gloria Isabel Ribera Martínez, and field technicians Leobardo González García, Octavia Pérez Medina, and Sandra Meza Morga for their valuable support in the field study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CRISP | Centro Regional de Investigación en Salud Pública |
| INSP | Instituto Nacional de Investigación en Salud Pública |
References
- Organización Panamericana de la Salud, «Dengue: datos y análisis». Acceded: 7 de diciembre de 2025. [En línea]. Disponible en: https://www.paho.org/es/arbo-portal/dengue-datos-analisis.
- Secretaría de Salud, «Panorama Epidemiológico de Dengue 2024», gob.mx. Acceded: 7 de diciembre de 2025. [En línea]. Disponible en: http://www.gob.mx/salud/documentos/panorama-epidemiologico-de-dengue-2024.
- L. P. Campbell, C. Luther, D. Moo-Llanes, J. M. Ramsey, R. Danis-Lozano, y A. T. Peterson, «Climate change influences on global distributions of dengue and chikungunya virus vectors», Philos. Trans. R. Soc. Lond. B. Biol. Sci., vol. 370, n.o 1665, p. 20140135, abr. 2015. [CrossRef]
- L. R. Bowman, S. Donegan, y P. J. McCall, «Is Dengue Vector Control Deficient in Effectiveness or Evidence?: Systematic Review and Meta-analysis», PLoS Negl. Trop. Dis., vol. 10, n.o 3, p. e0004551, mar. 2016. [CrossRef]
- K. Saavedra-Rodriguez et al., «Parallel evolution of vgsc mutations at domains IS6, IIS6 and IIIS6 in pyrethroid resistant Aedes aegypti from Mexico», Sci. Rep., vol. 8, n.o 1, p. 6747, abr. 2018. [CrossRef]
- F. Solis-Santoyo et al., «Insecticide resistance in Aedes aegypti from Tapachula, Mexico: Spatial variation and response to historical insecticide use», PLoS Negl. Trop. Dis., vol. 15, n.o 9, p. e0009746, sep. 2021. [CrossRef]
- A. J. Janich et al., «Permethrin Resistance Status and Associated Mechanisms in Aedes albopictus (Diptera: Culicidae) From Chiapas, Mexico», J. Med. Entomol., vol. 58, n.o 2, pp. 739-748, mar. 2021. [CrossRef]
- C. Baak-Baak et al., «Human blood as the only source of Aedes aegypti in churches from Merida, Yucatan, Mexico», J. Vector Borne Dis., vol. 55, n.o 1, p. 58, 2018. [CrossRef] [PubMed]
- I. Fernández-Salas, Biología y control de Aedes aegypti: Manual de operaciones., 2a. Universidad Autónoma de Nuevo León., 2009.
- J. E. García-Rejón et al., «Mosquito Infestation and Dengue Virus Infection in Aedes aegypti Females in Schools in Mérida, México», Am. Soc. Trop. Med. Hyg., vol. 84, n.o 3, pp. 489-496, mar. 2011. [CrossRef]
- A. C. Morrison et al., «Aedes aegypti (Diptera: Culicidae) production from non-residential sites in the Amazonian city of Iquitos, Peru», Ann. Trop. Med. Parasitol., vol. 100, n.o sup1, pp. 73-86, abr. 2006. [CrossRef]
- C. M. Baak-Baak et al., «Vacant Lots: Productive Sites for Aedes (Stegomyia ) aegypti (Diptera: Culicidae) in Mérida City, México», J. Med. Entomol., vol. 51, n.o 2, pp. 475-483, mar. 2014. [CrossRef]
- Instituto Nacional de Estadística, Geografía e Informática., «Compendio de información geográfica municipal 2010. Tapachula, Chiapas.», 2010.
- Comisión Mexicana de Ayuda a Refugiados, «Estadísticas Comisión Mexicana de Ayuda a Refugiados». Acceded: 3 de diciembre de 2025. [En línea]. Disponible en: https://www.gob.mx/comar/documentos/estadisticas-comar-2013-2017.
- E. A. Zarate-Nahon et al., «Aedes aegypti Mosquitoes at Nonresidential Sites Might be Related to Transmission of Dengue Virus in Monterrey, Northeastern Mexico», Southwest. Entomol., vol. 38, n.o 3, pp. 465-476, sep. 2013. [CrossRef]
- KMZ to KML Converter Online | MyGeodata Cloud». Acceded: 7 de agosto de 2025. [En línea]. Disponible en: https://mygeodata.cloud/converter/kmz-to-kml.
- G. Obra, E. Rebua, A. M. Hila, S. Resilva, R. Lees, y W. Mamai, «Ovitrap Monitoring of Aedes aegypti and Aedes albopictus in Two Selected Sites in Quezon City, Philippines», Philipp. J. Sci., vol. 151, n.o 5, ago. 2022. [CrossRef]
- C. Villarreal-Treviño, J. C. Rios Delgado, K. M. Valdez Delgado, J. A. Nettel-Cruz, y F. A. Zumaya Estrada, Manual de procedimientos para la cría de aedinos y anophelinos en condiciones de insectario. In Cría de mosquitos Culicidae y evaluación de insecticidasde uso en salud pública, 1a Edición. Danis Lozano R, Correa Morales F. Eds. Cuernavaca, Morelos, México: Instituto Nacional de Salud Pública, 2021.
- R. F. Darsie y R. A. Ward, Identification and geographical distribution of the mosquitoes of North America, north of Mexico. 2005.
- J. H. Zar, Biostatistical Analysis, 5th Edition. Pearson Prentice Hall, 2010.
- H. I. Sasmita et al., «Ovitrap surveillance of dengue vector mosquitoes in Bandung City, West Java Province, Indonesia», PLoS Negl. Trop. Dis., vol. 15, n.o 10, p. e0009896, oct. 2021. [CrossRef]
- I. C. Dos Reis, N. A. Honório, C. T. Codeço, M. D. A. F. M. Magalhães, R. Lourenço-de-Oliveira, y C. Barcellos, «Relevance of differentiating between residential and non-residential premises for surveillance and control of Aedes aegypti in Rio de Janeiro, Brazil», Acta Trop., vol. 114, n.o 1, pp. 37-43, abr. 2010. [CrossRef]
- I. Navarro-Kraul et al., «The Field Assessment of Quiescent Egg Populations of Aedes aegypti and Aedes albopictus during the Dry Season in Tapachula, Chiapas, Mexico, and Its Potential Impact on Vector Control Strategies», Insects, vol. 15, n.o 10, p. 798, oct. 2024. [CrossRef]
- R. Danis-Lozano, M. H. Rodriguez, L. Gonzalez-Ceron, y M. Hernandez-Avila, «Risk factors for Plasmodium vivax infection in the Lacandon forest, southern Mexico», Epidemiol. Infect., vol. 122, n.o 3, pp. 461-469, jun. 1999. [CrossRef]
Figure 1.
Geographic location and spatial distribution of ovitraps in Tapachula, Chiapas, Mexico. (A) Geographic location of Tapachula city within Chiapas state. (B) Distribution of ovitraps across residential and non-residential areas in Tapachula. (C) Close-up of ovitrap locations in the Laureles neighborhood. (D) Close-up of ovitrap locations in the public park “Los Cerritos”. (E) Close-up of ovitrap locations in the cemetery “Jardín”.
Figure 1.
Geographic location and spatial distribution of ovitraps in Tapachula, Chiapas, Mexico. (A) Geographic location of Tapachula city within Chiapas state. (B) Distribution of ovitraps across residential and non-residential areas in Tapachula. (C) Close-up of ovitrap locations in the Laureles neighborhood. (D) Close-up of ovitrap locations in the public park “Los Cerritos”. (E) Close-up of ovitrap locations in the cemetery “Jardín”.
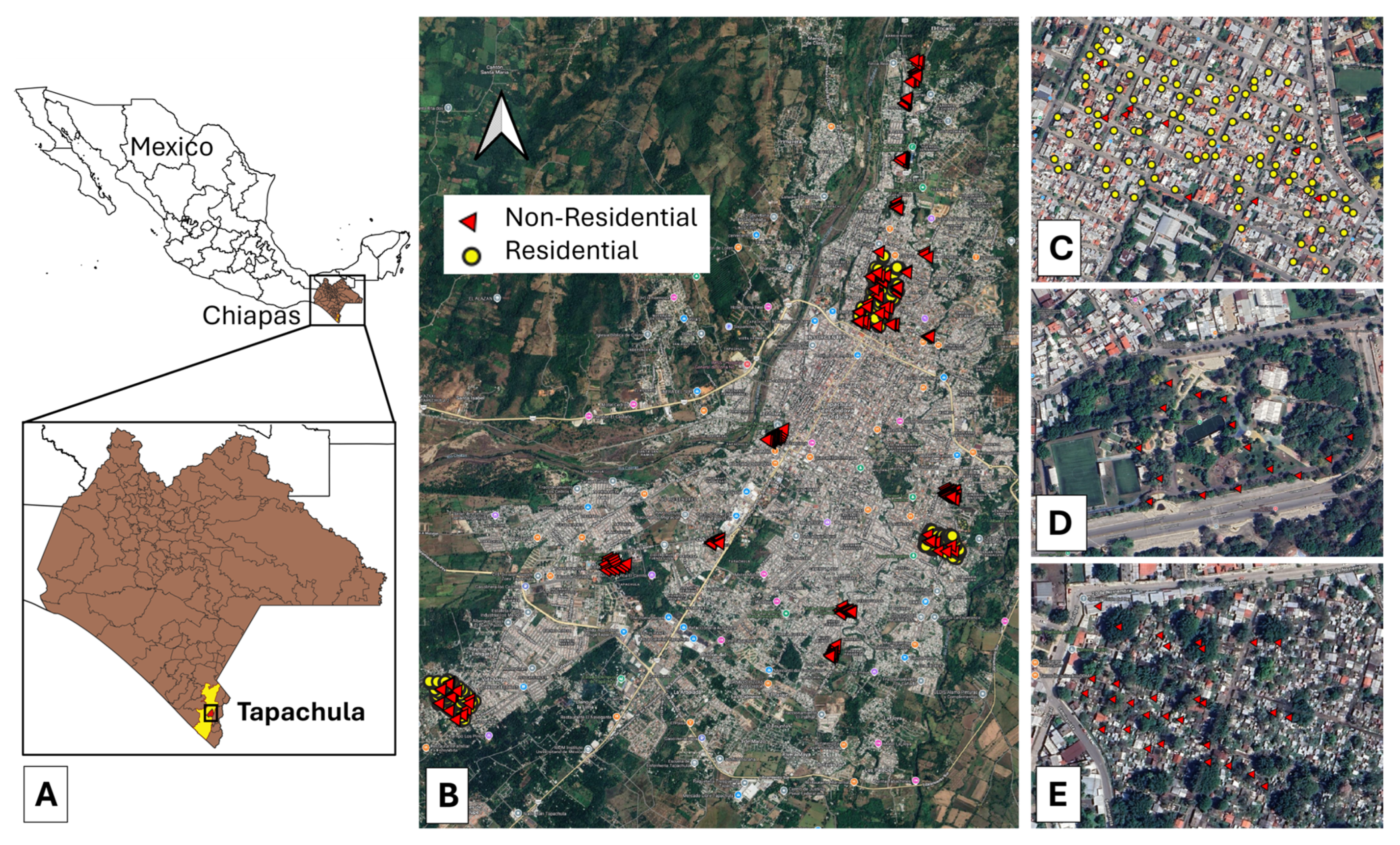
Figure 2.
Mean (±SE) number of eggs per ovitrap in residential and non-residential areas of Tapachula, Chiapas, Mexico.
Figure 2.
Mean (±SE) number of eggs per ovitrap in residential and non-residential areas of Tapachula, Chiapas, Mexico.
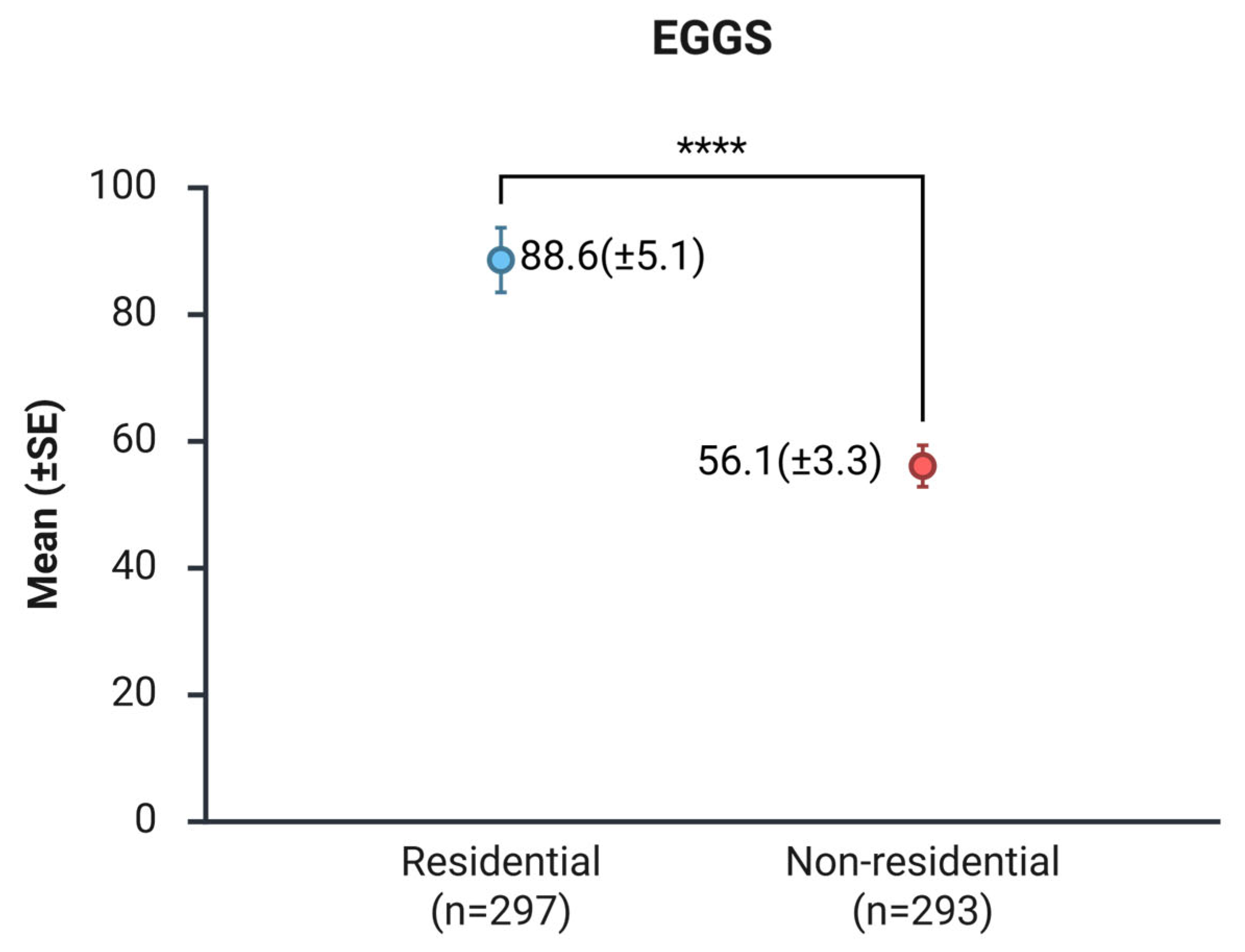
Figure 3.
Mean egg counts across various residential sites in Tapachula, Chiapas, Mexico.
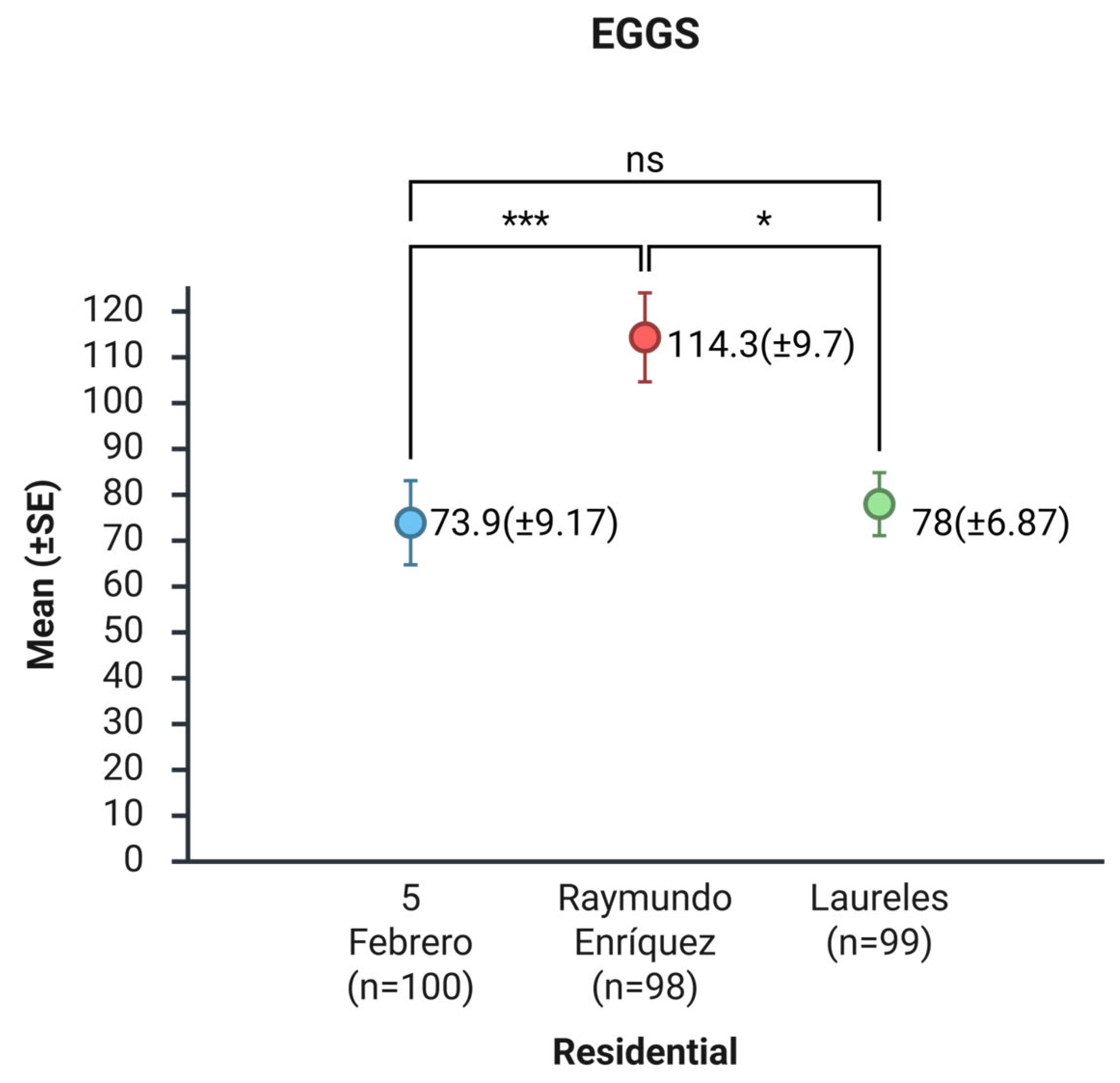
Figure 4.
Average egg counts across various non-residential site categories in Tapachula, Chiapas, Mexico.
Figure 4.
Average egg counts across various non-residential site categories in Tapachula, Chiapas, Mexico.
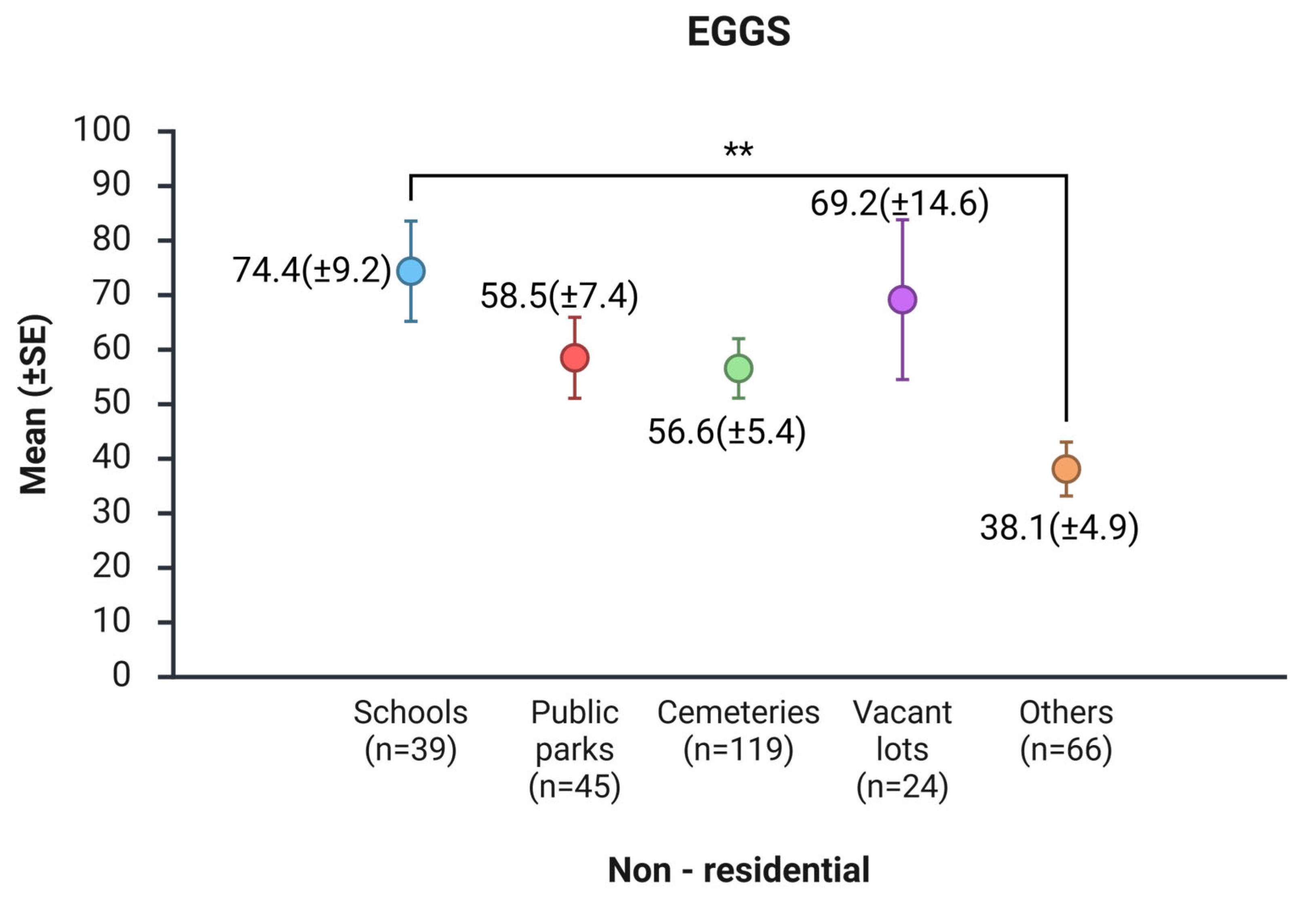
Figure 5.
Absolute (A) and relative (B) abundance of emerged Ae. aegypti and Ae. albopictus adults in residential and non-residential environments.
Figure 5.
Absolute (A) and relative (B) abundance of emerged Ae. aegypti and Ae. albopictus adults in residential and non-residential environments.
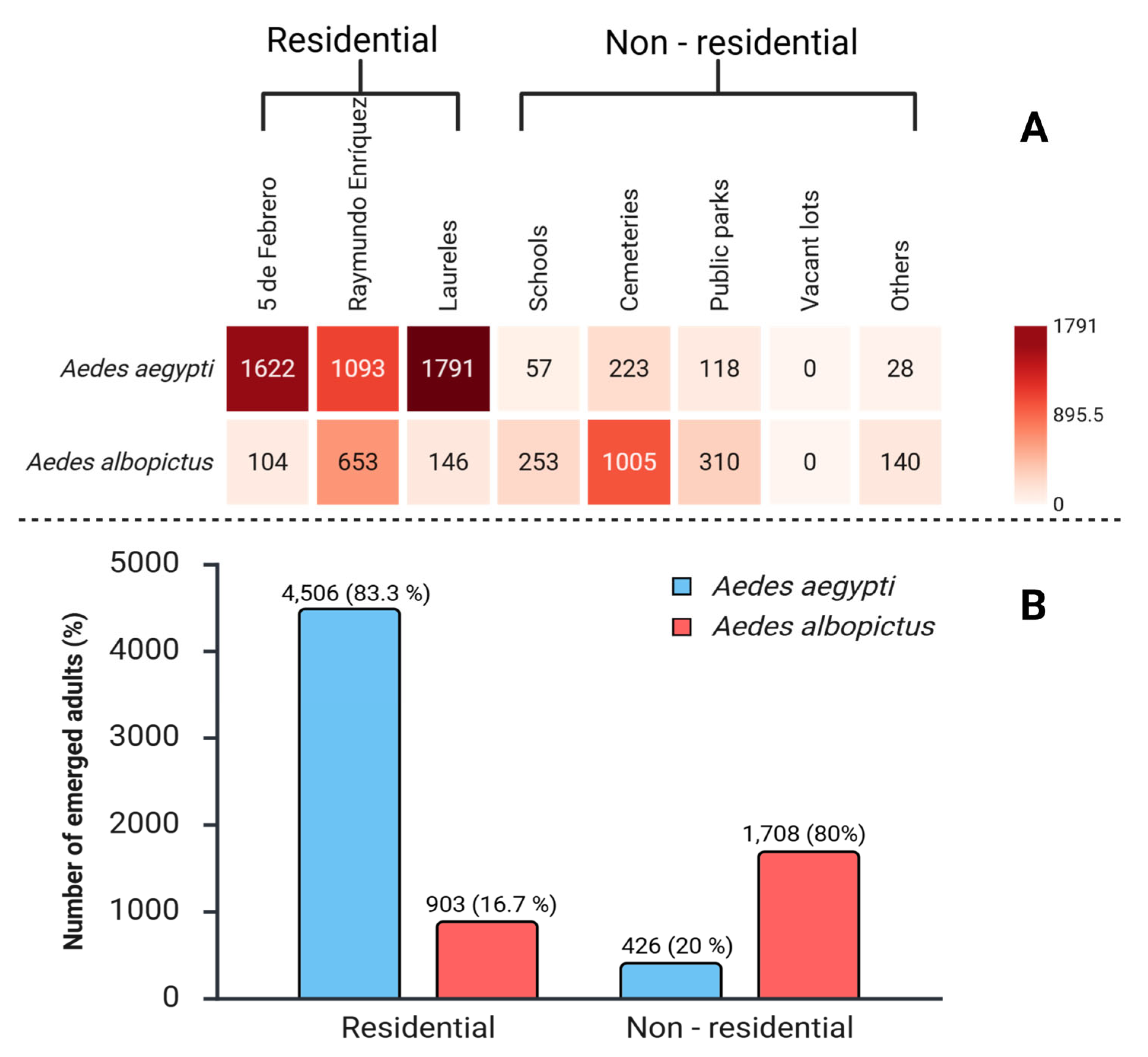
Figure 6.
Number of emerged adults from eggs among residential samples in Tapachula, Chiapas, Mexico.
Figure 6.
Number of emerged adults from eggs among residential samples in Tapachula, Chiapas, Mexico.
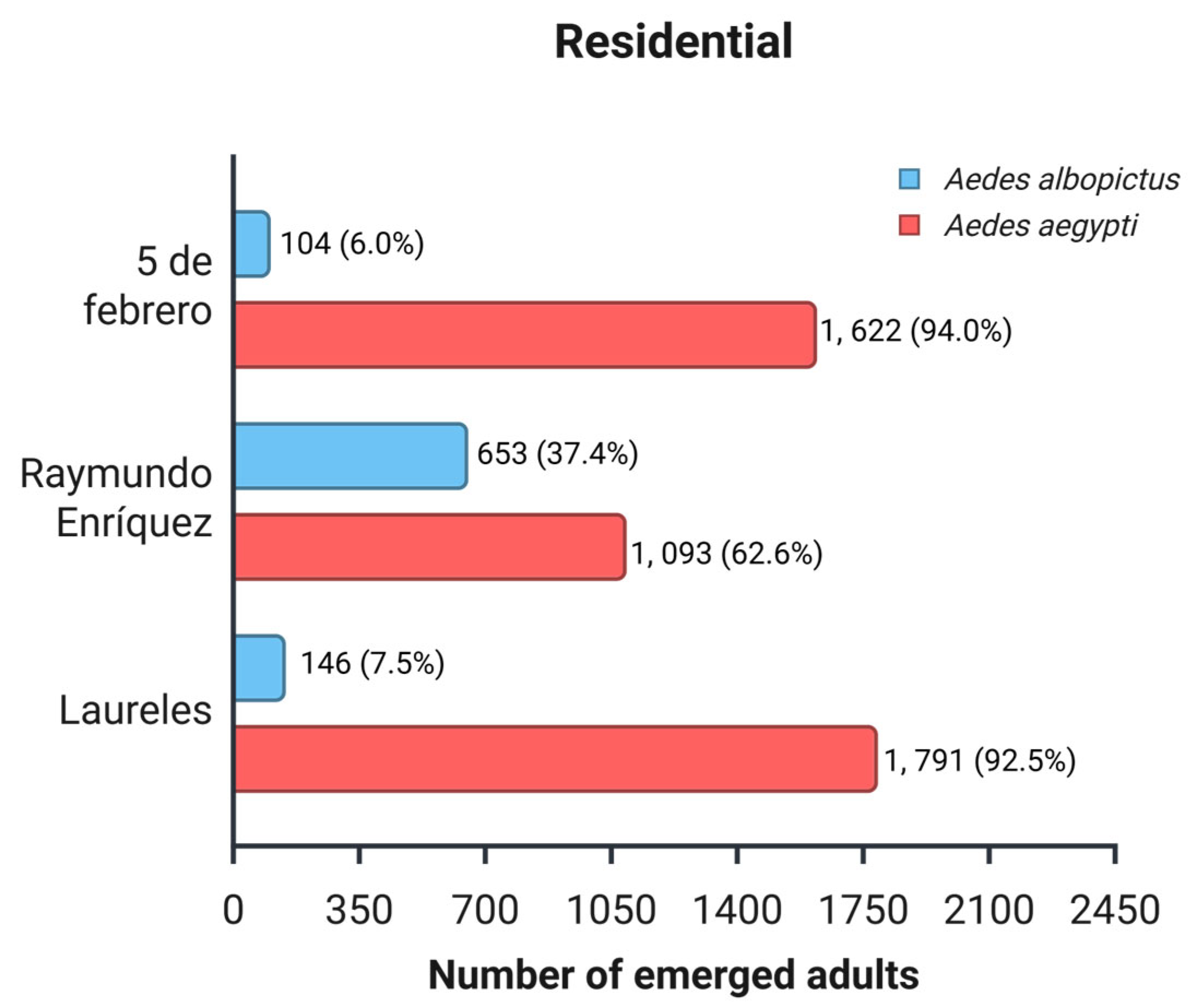
Figure 7.
Number of emerged adults from eggs among non-residential samples in Tapachula, Chiapas, Mexico.
Figure 7.
Number of emerged adults from eggs among non-residential samples in Tapachula, Chiapas, Mexico.
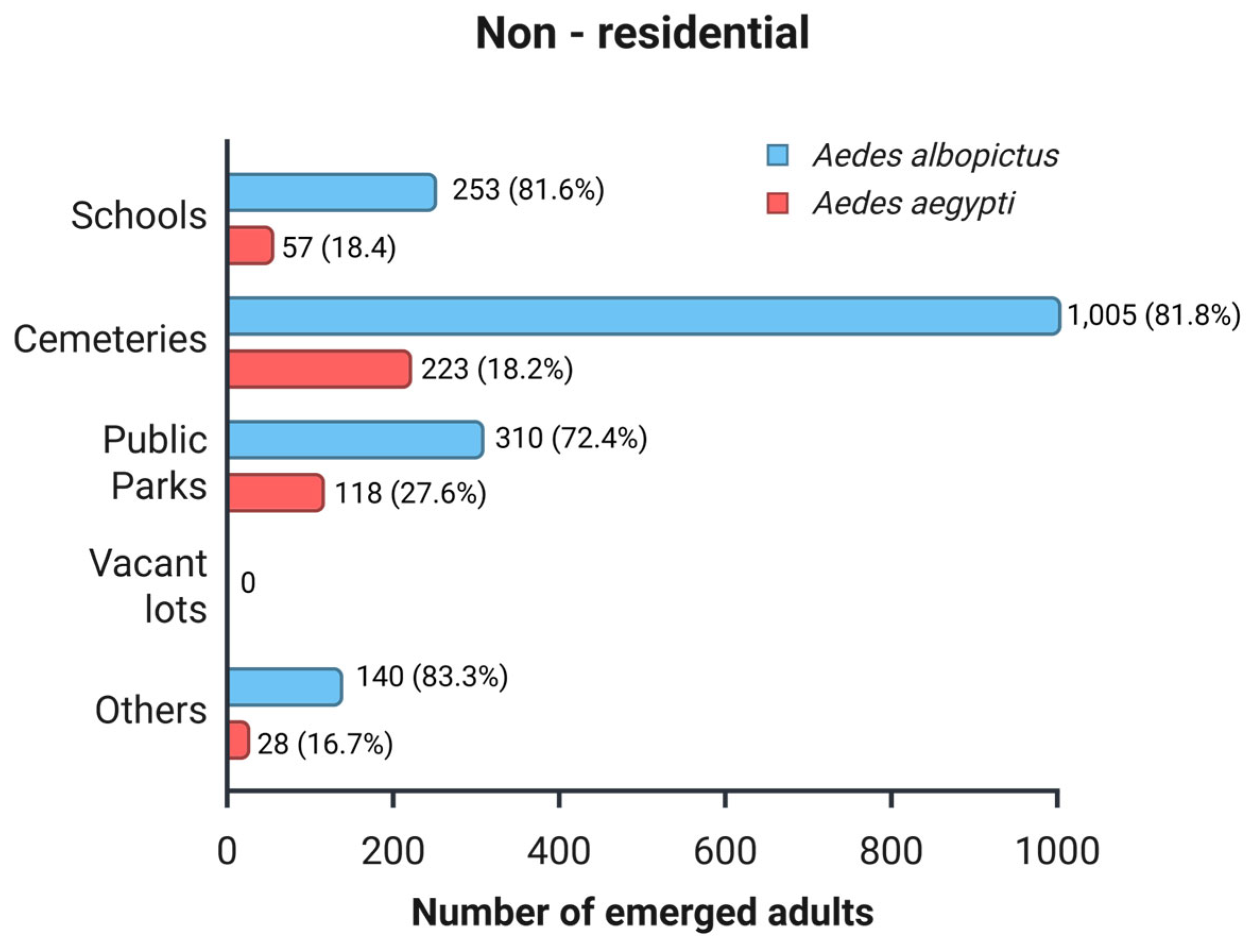

Table 1.
Ovitrap distribution by residential vs non-residential sites categories across sampled neighborhoods in Tapachula, Southern Mexico.
Table 1.
Ovitrap distribution by residential vs non-residential sites categories across sampled neighborhoods in Tapachula, Southern Mexico.
| Study area | Category | Sampled neighborhoods and categories of non-residential | ovitraps/ site (n) |
Total ovitraps (%) |
|---|---|---|---|---|
| Residential (R) | Neigborhood | 5 de Febrero | 100 | 297 (50) |
| Raymundo Enríquez | 98 | |||
| Laureles | 99 | |||
| Non-residential (NR) | Cemetery | Jardín | 30 | 119 (20) |
| Municipal | 30 | |||
| Indeco | 29 | |||
| Prados | 30 | |||
| Public Park | Los Cerritos | 15 | 45 (7.6) | |
| Indeco | 15 | |||
| Del Café | 15 | |||
| School | Preparatoria 4 | 4 | 39 (6.6) | |
| Teodomiro | 8 | |||
| Fray Matías | 4 | |||
| República de Cuba | 4 | |||
| 5 de febrero | 2 | |||
| Teófilo de Acebo | 6 | |||
| Técnica 3 | 4 | |||
| Raymundo Enríquez | 4 | |||
| Colinas del Rey | 3 | |||
| Vacant lots (1 ovitrap per vacant lots) | 26 | 26 (4.4) | ||
| Car junk lots | 5 de Febrero | 15 | 31 (5.2) | |
| Banorte | 16 | |||
| Mechanical workshop | MS1 | 2 | 29 (4.9) | |
| MS2 | 2 | |||
| MS3 | 2 | |||
| MS4 | 2 | |||
| MS5 | 1 | |||
| MS6 | 1 | |||
| MS7 | 1 | |||
| MS8 | 1 | |||
| MS9 | 1 | |||
| MS10 | 1 | |||
| MS11 | 3 | |||
| MS12 | 1 | |||
| MS14 | 1 | |||
| MS15 | 2 | |||
| MS16 | 2 | |||
| MS17 | 2 | |||
| MS18 | 2 | |||
| MS19 | 1 | |||
| MS20 | 1 | |||
| Car wash | CW1 | 3 | 5 (0.8) | |
| CW2 | 2 | |||
| Tire-repair shop | TRS1 | 2 | 2 (0.3) | |
| Total R | 297 (50) | |||
| Total NR | 293 (50) | |||
| Total R + NR | 593 (100) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



