
Submitted:
25 August 2025
Posted:
26 August 2025
You are already at the latest version
Abstract
Perfluorooctane sulfonamide (PFOSA), the direct precursor to perfluorooctane sul-fonate (PFOS), is widely presented in the environment. Research has indicated that PFOSA is cardiotoxic and hepatotoxic, but its impact on neurodevelopment remains unclear. In the current study, we observed that exposure of PFOSA caused neurode-velopmental toxicity in zebrafish embryos in a dose-dependent manner, as evidenced by impaired motor abilities and decreased swimming distance. We then demonstrated that PFOSA exposure downregulated the mRNA expression of neurodevelopment-related genes including a1-tubulin, elavl3, ache and dat. Moreover, PFOSA exposure resulted in dose-dependent oxidative stress, which triggers apoptosis in the brains of zebrafish larvae. We further showed that inhibition of the aryl hydrocarbon receptor (AhR) alle-viated the oxidative stress and apoptosis induced by PFOSA-induced, thereby coun-teracting the neurodevelopmental abnormalities in zebrafish larvae. In conclusion, these findings indicate PFOSA causes neurodevelopmental disorders by inducing oxi-dative stress and apoptosis through the AhR pathway.
Keywords:
PFOSA
; AhR
; oxidative stress
; apoptosis
; neurotoxicity
1. Introduction
Per- and polyfluoroalkyl substances (PFAS) constitute a class of persistent organic contaminants that have raised significant public health concerns due to their widespread presence in environment and their tendency to bioaccumulation [1]. Perfluorooctane sulfonamide (PFOSA), the immediate precursor of perfluorooctane sulfonic acid (PFOS), is frequently detected in soil, surface water, and groundwater [2,3]. The concentrations of PFOSA can be up to 15 μg/L in surface waters and 0.09-20,000 μg/kg in surface-soil [2,4]. Notably, PFOSA has been reported as the only PFOS-related precursor (PreFOS) detected in all water and sediment samples from Taihu Lake, China [5]. PFOSA was also found to be the most abundant PreFOS in all fish tissues from Taihu Lake [6]. Moreover, the concentrations of PFOSA in tissues of finless porpoises from East China Sea are increasing with time between 2009-2010 and 2018-2019 [7].
Humans are primarily exposure to PFOSA through the ingestion of contaminated food and water, with concentrations in human blood reaching up to 1.6 μg/L [8]. Importantly, PFOSA can cross the placental barrier, posing potential health risks to developing fetuses [9]. Among 38 tested PFAS compounds, PFOSA was uniquely reported to cause embryonic toxicity in zebrafish at low concentrations [10]. Chen et al reported that PFOSA, at concentrations ranging from 0.1-100 μg/L significantly reduced heartbeat rate, stroke volume, and cardiac output in zebrafish [11]. Additionally, exposure to PFOSA can lead to liver and kidney damage in zebrafish embryos [12,13]. Research on the neurotoxic effects of PFOSA is limited. Slotkin et al. reported that PFOSA has a more detrimental impact on the rat neural cell line PC12 compared to PFOS, perfluorooctanoic acid (PFOA), and perfluorobutane sulfonate (PFBS) [14]. A recent study also demonstrated that PFOSA exposure impaired neurodevelopment in zebrafish embryos in a dose-dependent manner [15]. However, the specific mechanisms behind PFOSA's neurodevelopmental toxicity remain unclear.
While most PFASs act as agonists of peroxisome proliferator-activated receptors (PPARs), PFOSA predominantly activates the aryl hydrocarbon receptor (AhR) signaling pathway [11,16]. AhR is a ligand-activated transcription factor participate in multiple cellular processes. Activated AhR translocates to the nucleus, where it binds to xenobiotic response element (XRE) sites in the promoter regions of target genes, regulating their transcription [17]. It has been reported that the AhR signaling pathway mediated 3,6-dibromocarbazole (3,6-DBCZ)-induced neurodevelopmental toxicity in juvenile zebrafish [18]. The AhR target genes, such as cytochrome P450 CYP1s, can generate reactive oxygen species (ROS) as a byproduct during xenobiotic metabolism [19]. Elevated ROS levels can induce oxidative damage, impair neural structure and function, and lead to neuronal injury or death [20].
Apoptosis, also known as programmed cell death, can be classified into two types: intrinsic apoptosis, which is mediated by mitochondria, and extrinsic apoptosis, which is mediated by death receptors. It is widely acknowledged that apoptosis is crucial for neurodevelopment, particularly in shaping the developing brain [21]. However, excessive apoptosis is often a significant factor in developmental neurotoxicity [22]. Numerous environmental chemicals with neurodevelopmental toxicity, such as metals, pesticides, and endocrine-disrupting compounds, can trigger neuronal cell apoptosis [22]. Our recent findings indicate that exposure to PFOSA leads to apoptosis through AhR-mediated oxidative stress in the hearts of zebrafish embryos [16]. Consequently, we propose that PFOSA induces apoptosis and disrupts neurodevelopment through the AhR/ROS axis.
The zebrafish (Danio rerio) model offers distinct advantages for assessing developmental neurotoxicity, including genetic tractability, optical transparency, and conserved neurotransmitter systems [23]. Zebrafish behavioral phenotypes have emerged as sensitive biomarkers for neurotoxicant screening, with locomotor deficits strongly correlating with neurodevelopmental outcomes in mammals [23,24]. In this study, we characterize PFOSA-induced neurodevelopmental toxicity in zebrafish embryos and explored the role of the AhR /ROS axis in this process.
2. Materials and Methods
2.1. Chemicals
PFOSA (CAS: 754-91-6, purity 94.9%) was obtained from Scrbio, China. The AhR inhibitor CH223191(CH, CAS:301326-22-7, purity 99.64%) and the ROS scavenger NAC (CAS: 616-91-1, purity >98%) were purchased from AbMole, Shanghai China and Beyotime, Shanghai, China, respectively. All these chemicals were dissolved in dimethyl sulfoxide (DMSO) to obtain a stock solution and stored in -80 °C.
2.2. Zebrafish Husbandry and Chemical Exposure
Adult wild-type AB strain zebrafish were kept at 28.5 °C under a 14:10 h light: dark cycle in a recirculating aquaculture setup with E3 medium consisting of 5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, and 0.33 mM MgSO4. Brine shrimp were provided as nourishment to the fish twice daily. Spawning pairs, consisting of two female and one male zebrafish, were transferred to breeding tanks, and fertilized eggs were collected the following day. At 2 h post-fertilization (hpf), embryos were treated with PFOSA at different concentrations (6.25, 12.5, 25, 50 μg /L) in the presence or absence of CH (0.05 μM) or NAC (0.25µM) until 72 hpf. DMSO (0.01 %, v/v) served as the vehicle control. Exposure solutions were refreshed every 24 h.
2.3. Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Total RNA was isolated from the head of zebrafish larvae using Trizol reagent (Vazyme, Nanjing, China). RNA purity and concentration were evaluated by using a NanoDrop 2000 spectrophotometer (NanoDrop Technology, DE, U.S.). complementary DNA (cDNA) was synthesized using a commercial reverse transcription kit (Vazyme, Nanjing, China). RT-PCR amplifications were conducted on an ABI 7500 real-time-PCR system (Applied Biosystems, CA, U.S.) using SYBR Green PCR Master Mix (Vazyme, Nanjing, China). Primer sequences are listed in Table 1. The thermal cycling conditions included an initial denaturation for 2 minutes at 50 °C, followed by 10 minutes at 95 °C, then 40 cycles of 95 °C for 15 s and annealing at 60 °C for 60 s. Relative gene expression levels were normalized to β-Actin and calculated using the 2-△△CT method.
2.4. Zebrafish Behavioral Tests
Locomotion activity was assessed as reported previously [25]. Briefly, at 72 hpf, zebrafish embryos from each group were individually placed to wells of a 48-well plate filled with 1 ml of E3 medium. After a 10-minute acclimation period, their movements were recorded during two light-dark circles. The locomotor behavior was analyzed using Danio Vision (Noldus, the Netherlands), and parameters such as cumulative movement duration, total swimming distance, and mean velocity were measured and analyzed with EthoVision XT 15 software (Noldus, the Netherlands). All trials were independently replicated in triplicate.
2.5. ROS Detection
ROS levels were assessed using DCFH-DA (2',7'-Dichlorodihydrofluorescein diacetate) staining. Ten embryos at 72 hpf from each group were incubated with 5 μg/mL DCFH-DA in darkness for 30 minutes. Following washing with PBS thrice, fluorescence images were captured under a fluorescence microscope.
2.6. Detection of Apoptosis
Acridine orange (AO) staining was utilized to assess apoptosis levels in the brain region of zebrafish larvae. Ten zebrafish embryos from each group were immersed in acridine orange solution (5 mg/L) in darkness for 30 minutes. After washing three times with PBS, larvae were observed under a fluorescence microscope (Nikon, SMZ18). Images were analyzed with ImageJ software.
2.7. Statistical Analysis
All experiments were replicated at least three times under independent conditions. Statistical significance was performed using one-way ANOVA followed by Dunnett’s or Tukey’s multiple comparison tests, as appropriate. Data are presented as mean ± SEM (Standard Error of the Mean). A P values <0.05 was deemed statistically significant.
3. Results
3.1. PFOSA Exposure Induces Locomotor Deficits and Behavioral Preference Alterations and in Zebrafish Larvae.
As shown in Figure 1A,B, no significant differences in survival rates or hatching rates were observed in zebrafish embryos exposed to PFOSA at different concentrations (0-50 μg/L) when compared to DMSO controls. However, behavioral analysis revealed that PFOSA exposure led to a dose-dependent reduction in the complexity of movement trajectories (Figure 1C). Moreover, exposure to PFOSA significantly reduced total swimming distance and decreased swimming speed at concentrations exceeding 6.25 µg/L (Figure 1D).
3.2. PFOSA Exposure Impairs Neuronal Differentiation
As illustrated in Figure 2, PFOSA exposure downregulated the mRNA expression levels of α1-tubulin (an early neuronal differentiation marker) and elavl3 (a marker of mature neurons) in a dose dependent manner. Notably, PFOSA even at the lowest tested concentration of 6.25 μg/L significantly decreased the transcriptional levels of these two gene. Additionally, PFOSA at concentrations above 6.25 μg/L significantly decreased the mRNA levels of ache and dat (acetylcholinesterase and dopamine transporter, both are key neurotransmitter system components).
3.3. PFOSA Exposure Leads to Oxidative Stress and Apoptosis in Zebrafish Brains
PFOSA at dose levels above 12.5 induced a concentration-dependent increase in ROS production in embryonic heads (Figure 3A). We further demonstrated that PFOSA induced apoptosis in a concentration-dependent manner, evidenced by increased apoptotic bodies in the head of zebrafish larvae (Figure 3B). In consistent, the mRNA expression levels of the oxidative stress-related gene sod2 and the pro-apoptotic gene tp53 were elevated in the groups exposed to PFOSA at 25 and 50 µg/L (Figure 3C). The mRNA levels of nrf2a, sod1 and bax were also significantly increased in the group with the highest concentration of PFOSA (Figure 3C)
3.4. PFOSA Triggers Apoptosis via AhR-Mediated Oxidative Stress
We first observed that the mRNA levels of cyp1a1 and ahrra, two prototypical AhR downstream genes, were elevated in high concentration (50 µg/L) PFOSA samples but returned to normal levels in the presence of CH, indicating that AhR signaling was activated by PFOSA (Figure 4A). We then demonstrated that the addition of the AhR inhibitor CH as well as the ROS scavenger NAC counteracted oxidative stress and apoptosis in the brain region of zebrafish larvae exposed to PFOSA (Figure 4B,C). Notably, co-treatment with CH abolished the behavioral deficits in locomotor activity caused by PFOSA (Figure 4D). The aberrant expression patterns of neuro-differentiation genes (α1-tubulin, elavl3, ache and dat) in PFOSA samples were also returned to control levels in the PFOSA plus CH group (Figure 4E).
4. Discussion
Behavioral analysis is recognized as an effective approach for evaluating neurotoxicity [26]. In this study, we observed that zebrafish embryos exposure to PFOSA resulted in a dose-dependent reduction in the complexity of movement trajectories, total swimming distance, and swimming speed. Moreover, the mRNA expression levels of neurodevelopmental related genes—including α1-tubulin, elavl3, ache and dat —were downregulated in the head of zebrafish embryos exposed to PFOSA. These findings are in line with a recent study that reported PFOSA exposure reduced locomotor activity of larval fish and decreased the expression levels of elavl3 and ache [15]. α1-tubulin is an essential component of the microtubule cytoskeleton and plays a key role in the structural and functional integrity of axons and dendrites [27]. Elavl, a neural-specific RNA-binding protein, is involved in neurogenesis [28]. AChE activity is crucial for the inactivation of acetylcholine at nerve terminals and for the proper functioning of the sensory and neuromuscular systems [29]. Dat as a dopamine transporter regulates synaptic dopamine levels, which are vital for social behavior in fish [30]. The downregulation of these genes suggests that PFOSA exposure not only impacts neurogenesis but also disrupted the structure and function of nervous system.
Neural development relies on tightly regulation of neurogenesis and apoptosis [31]. In this study, we observed that PFOSA exposure induced a dose-dependent increase in apoptotic bodies in the heads of zebrafish larvae. Additionally, the mRNA levels of pro-apoptotic genes including tp53 and bax were elevated following PFOSA exposure. The transcription factor p53, encoded by tp53, plays a key role in cellular stress responses. Activated p53 can elicit apoptosis in various cell types including neurons [32]. Bax, a member of the Bcl-2 family, is a direct target of p53. Activation of Bax by p53 can cause mitochondria permeabilization and initiate intrinsic apoptosis [33]. The increase in apoptotic bodies and the upregulation of tp53 and bax have been reported in the hearts of zebrafish embryos exposed to PFOSA, suggesting that PFOSA might trigger apoptosis through a similar mechanism across different organs [16].
Oxidative stress arises when there is an imbalance between the generation of ROS and the body’s capability to neutralize them, often resulting in cellular damage and death [34]. It has been established that oxidative stress, as a main driver of apoptosis, is a critical factor in neurodevelopmental toxicity [35]. We recently reported that PFOSA exposure induced oxidative stress and apoptosis in zebrafish embryonic hearts [16]. In this study, we observed a dose-dependent elevation in ROS production and the upregulation of oxidative related genes, including nrf2, sod1, and sod2, in the head of zebrafish larvae exposed to PFOSA. NAC is among the most widely used antioxidant agents [36]. The addition of ROS scavenger NAC not only mitigated oxidative stress but also diminished apoptosis in the brain area of zebrafish larvae exposed to PFOSA. Nrf2 serves as a the key transcription factor in the antioxidant defense system, while Sod1 and Sod2 are crucial for detoxifying superoxide radicals [37]. Our findings suggest that antioxidant treatment could be a promising therapeutic strategy to alleviate PFOSA-induced damage during neural development.
AhR, which is expressed in the vertebrate brain in the early developmental stages, plays an essential role in neurogenesis [38,39]. Overactivation of AhR has been shown to promote neuronal cell apoptosis in the hippocampus of mice [40]. Recent studies showed that AhR is involved in PFOSA-caused cardiac defects in zebrafish embryos [11,16]. Here, we demonstrated that PFOSA activated AhR in the head of zebrafish larvae, evidenced by the overexpression of downstream genes cyp1a1 and ahrra. Cyp1a1, a prototypical AhR target gene, is important not only for xenobiotic metabolism, but also plays a key role in AhR-induced oxidative stress [41]. We have previously reported that AhR activation by benzo[a]pyrene leads to ROS overproduction through Cyp1a1 in zebrafish [42]. There are two Ahrr isoforms in zebrafish, with Ahrra primarily responsible for regulating AHR signaling during development [43]. In consistent, we observed that inhibition of AhR diminished oxidative stress, apoptosis and behavior abnormalities caused by PFOSA exposure.
5. Conclusions
In summary, our results indicate that PFOSA exposure elicits oxidative stress and promotes apoptosis in the brains of zebrafish embryos via AhR activation, leading to abnormal neurodevelopment. Our study contributes to understanding the molecular mechanisms underlying the neurodevelopmental toxicity of PFOSA, highlighting the ecological and health risks of this persistent pollutant. Further studies utilizing mammalian models are necessary to evaluate the potential risks of PFOSA to human health.
Author Contributions
Investigation and writing—original draft preparation, PY.C.; investigation, K.W.; resources, J.Z.; funding acquisition, supervision, writing—review and editing, Y.J; project administration, supervision, writing—review and editing, T.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant number: 81870239) and The Priority Academic Program Development of Jiangsu Higher Education Institutions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ateia, M. and M. Scheringer, From "forever chemicals" to fluorine-free alternatives. Science, 2024. 385(6706): p. 256-258.
- Brusseau, M.L.; Anderson, R.H.; Guo, B. PFAS concentrations in soils: Background levels versus contaminated sites. Sci. Total. Environ. 2020, 740, 140017–140017. [Google Scholar] [CrossRef]
- Anderson, R.H.; Long, G.C.; Porter, R.C.; Anderson, J.K. Occurrence of select perfluoroalkyl substances at U.S. Air Force aqueous film-forming foam release sites other than fire-training areas: Field-validation of critical fate and transport properties. Chemosphere 2016, 150, 678–685. [Google Scholar] [CrossRef]
- Anderson, R.H.; Long, G.C.; Porter, R.C.; Anderson, J.K. Occurrence of select perfluoroalkyl substances at U.S. Air Force aqueous film-forming foam release sites other than fire-training areas: Field-validation of critical fate and transport properties. Chemosphere 2016, 150, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, Q.; Shan, G.; Zhu, L.; Yang, L.; Liu, M. Occurrence, partitioning and bioaccumulation of emerging and legacy per- and polyfluoroalkyl substances in Taihu Lake, China. Sci. Total. Environ. 2018, 634, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhu, L.; Wang, Q.; Shan, G. Tissue distribution and bioaccumulation of legacy and emerging per-and polyfluoroalkyl substances (PFASs) in edible fishes from Taihu Lake, China. Environ. Pollut. 2021, 268, 115887. [Google Scholar] [CrossRef]
- Zhang, B.; He, Y.; Yang, G.; Chen, B.; Yao, Y.; Sun, H.; Kannan, K.; Zhang, T. Legacy and Emerging Poly- and Perfluoroalkyl Substances in Finless Porpoises from East China Sea: Temporal Trends and Tissue-Specific Accumulation. Environ. Sci. Technol. 2021, 56, 6113–6122. [Google Scholar] [CrossRef]
- Ding, J.; Dai, Y.; Zhang, J.; Wang, Z.; Zhang, L.; Xu, S.; Tan, R.; Guo, J.; Qi, X.; Chang, X.; et al. Associations of perfluoroalkyl substances with adipocytokines in umbilical cord serum: A mixtures approach. Environ. Res. 2022, 216. [Google Scholar] [CrossRef]
- Liu, L.; Yan, P.; Liu, X.; Zhao, J.; Tian, M.; Huang, Q.; Yan, J.; Tong, Z.; Zhang, Y.; Zhang, J.; et al. Profiles and transplacental transfer of per- and polyfluoroalkyl substances in maternal and umbilical cord blood: A birth cohort study in Zhoushan, Zhejiang Province, China. J. Hazard. Mater. 2024, 466, 133501. [Google Scholar] [CrossRef]
- Dasgupta, S.; Reddam, A.; Liu, Z.; Liu, J.; Volz, D.C. High-content screening in zebrafish identifies perfluorooctanesulfonamide as a potent developmental toxicant. Environ. Pollut. 2020, 256, 113550–113550. [Google Scholar] [CrossRef]
- Chen, H.; Qiu, W.; Yang, X.; Chen, F.; Chen, J.; Tang, L.; Zhong, H.; Magnuson, J.T.; Zheng, C.; Xu, E.G. Perfluorooctane Sulfonamide (PFOSA) Induces Cardiotoxicity via Aryl Hydrocarbon Receptor Activation in Zebrafish. Environ. Sci. Technol. 2022, 56, 8438–8448. [Google Scholar] [CrossRef]
- Dasgupta, S.; Reddam, A.; Liu, Z.; Liu, J.; Volz, D.C. High-content screening in zebrafish identifies perfluorooctanesulfonamide as a potent developmental toxicant. Environ. Pollut. 2020, 256, 113550–113550. [Google Scholar] [CrossRef] [PubMed]
- Xuan, R.; Qiu, X.; Wang, J.; Liu, S.; Magnuson, J.T.; Xu, B.; Qiu, W.; Zheng, C. Hepatotoxic response of perfluorooctane sulfonamide (PFOSA) in early life stage zebrafish (Danio rerio) is greater than perfluorooctane sulfonate (PFOS). J. Hazard. Mater. 2023, 461, 132552. [Google Scholar] [CrossRef]
- Slotkin, T.A.; MacKillop, E.A.; Melnick, R.L.; Thayer, K.A.; Seidler, F.J. Developmental Neurotoxicity of Perfluorinated Chemicals Modeled in Vitro. Environ. Heal. Perspect. 2008, 116, 716–722. [Google Scholar] [CrossRef]
- David, N.; Ivantsova, E.; Konig, I.; English, C.D.; Avidan, L.; Kreychman, M.; Rivera, M.L.; Escobar, C.; Valle, E.M.A.; Sultan, A.; et al. Adverse Outcomes Following Exposure to Perfluorooctanesulfonamide (PFOSA) in Larval Zebrafish (Danio rerio): A Neurotoxic and Behavioral Perspective. Toxics 2024, 12, 723. [Google Scholar] [CrossRef]
- Ma, T.; Jiang, Y.; Chen, P.; Xiao, F.; Zhang, J.; Ma, Y.; Chen, T. PFOS and PFOSA induce oxidative stress-mediated cardiac defects in zebrafish via PPARγ and AHR pathways, respectively. Sci. Total. Environ. 2024, 951, 175716. [Google Scholar] [CrossRef]
- Polonio, C.M.; McHale, K.A.; Sherr, D.H.; Rubenstein, D.; Quintana, F.J. The aryl hydrocarbon receptor: a rehabilitated target for therapeutic immune modulation. Nat. Rev. Drug Discov. 2025, 24, 610–630. [Google Scholar] [CrossRef]
- Wang, X.; Hu, M.; Li, M.; Huan, F.; Gao, R.; Wang, J. Effects of exposure to 3,6-DBCZ on neurotoxicity and AhR pathway during early life stages of zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2023, 270, 115892. [Google Scholar] [CrossRef]
- Veith, A.; Moorthy, B. Role of cytochrome P450s in the generation and metabolism of reactive oxygen species. Curr. Opin. Toxicol. 2018, 7, 44–51. [Google Scholar] [CrossRef]
- Wu, Y.; Fan, F.; Zhou, L.; Shen, Y.; Wang, A.; Qin, Y.; Wang, J.; Yao, W. ADB-FUBINACA-induced developmental toxicity, neurotoxicity, and cardiotoxicity in embryonic zebrafish (Danio rerio). Environ. Res. 2025, 276, 121517. [Google Scholar] [CrossRef]
- Lein, P.J., S. Supasai, and M. Guignet, Chapter 9 - Apoptosis as a Mechanism of Developmental Neurotoxicity, in Handbook of Developmental Neurotoxicology (Second Edition), W. Slikker, M.G. Paule, and C. Wang, Editors. 2018, Academic Press. p. 91-112.
- Rock, K.D.; Patisaul, H.B. Environmental Mechanisms of Neurodevelopmental Toxicity. Curr. Environ. Heal. Rep. 2018, 5, 145–157. [Google Scholar] [CrossRef] [PubMed]
- de Abreu, M.S., et al., Zebrafish as a Model of Neurodevelopmental Disorders. Neuroscience, 2020. 445: p. 3-11.
- Vaz, R.; Hofmeister, W.; Lindstrand, A. Zebrafish Models of Neurodevelopmental Disorders: Limitations and Benefits of Current Tools and Techniques. Int. J. Mol. Sci. 2019, 20, 1296. [Google Scholar] [CrossRef]
- Nery, L.R.; Eltz, N.S.; Hackman, C.; Fonseca, R.; Altenhofen, S.; Guerra, H.N.; Freitas, V.M.; Bonan, C.D.; Vianna, M.R.M.R.; Laks, J. Brain Intraventricular Injection of Amyloid-β in Zebrafish Embryo Impairs Cognition and Increases Tau Phosphorylation, Effects Reversed by Lithium. PLOS ONE 2014, 9, e105862. [Google Scholar] [CrossRef]
- Barreiros, M.d.O.; Barbosa, F.G.; Dantas, D.d.O.; Santos, D.d.M.L.d.; Ribeiro, S.; Santos, G.C.d.O.; Barros, A.K. Zebrafish automatic monitoring system for conditioning and behavioral analysis. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.-Y.; Wu, Y.-Y.; Xia, B.; Dai, M.-Z.; Huang, Y.-F.; Yang, H.; Li, C.-Q.; Li, P. Fenobucarb-induced developmental neurotoxicity and mechanisms in zebrafish. NeuroToxicology 2020, 79, 11–19. [Google Scholar] [CrossRef]
- Fan, C.-Y.; Cowden, J.; Simmons, S.O.; Padilla, S.; Ramabhadran, R. Gene expression changes in developing zebrafish as potential markers for rapid developmental neurotoxicity screening. Neurotoxicology Teratol. 2010, 32, 91–98. [Google Scholar] [CrossRef]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a Biomarker in Environmental and Occupational Medicine: New Insights and Future Perspectives. BioMed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Du, Y.; Li, Q.; Zhou, G.; Cai, Z.; Man, Q.; Wang, W.C. Early-life perfluorooctanoic acid exposure disrupts the function of dopamine transporter protein with glycosylation changes implicating the links between decreased dopamine levels and disruptive behaviors in larval zebrafish. Sci. Total. Environ. 2024, 917, 170408. [Google Scholar] [CrossRef]
- van Leyen, K., et al., Neurogenesis and Apoptotic Cell Death. The Cell Cycle in the Central Nervous System, ed. D. Janigro. Vol. NJ. p. 2006, Humana Press: Totowa, . 71-79.
- Amaral, J.D.; Xavier, J.M.; Steer, C.J.; Rodrigues, C.M. The role of p53 in apoptosis. . 2010, 9, 145–52. [Google Scholar]
- Chipuk, J.E.; Kuwana, T.; Bouchier-Hayes, L.; Droin, N.M.; Newmeyer, D.D.; Schuler, M.; Green, D.R. Direct Activation of Bax by p53 Mediates Mitochondrial Membrane Permeabilization and Apoptosis. Science 2004, 303, 1010–1014. [Google Scholar] [CrossRef]
- Love, C.; Sominsky, L.; O’hEly, M.; Berk, M.; Vuillermin, P.; Dawson, S.L. Prenatal environmental risk factors for autism spectrum disorder and their potential mechanisms. BMC Med. 2024, 22, 1–13. [Google Scholar] [CrossRef]
- Nishimura, Y.; Kanda, Y.; Sone, H.; Aoyama, H.; Gasparovic, A.C. Oxidative Stress as a Common Key Event in Developmental Neurotoxicity. Oxidative Med. Cell. Longev. 2021, 2021, 6685204. [Google Scholar] [CrossRef]
- Kerksick, C.; Willoughby, D. The Antioxidant Role of Glutathione and N-Acetyl-Cysteine Supplements and Exercise-Induced Oxidative Stress. J. Int. Soc. Sports Nutr. 2005, 2, 38–44. [Google Scholar] [CrossRef]
- Giudice, A.; Montella, M. Activation of the Nrf2–ARE signaling pathway: a promising strategy in cancer prevention. BioEssays 2006, 28, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Latchney, S.E.; Hein, A.M.; O'BAnion, M.K.; DiCicco-Bloom, E.; Opanashuk, L.A. Deletion or activation of the aryl hydrocarbon receptor alters adult hippocampal neurogenesis and contextual fear memory. J. Neurochem. 2012, 125, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Juricek, L.; Coumoul, X. The Aryl Hydrocarbon Receptor and the Nervous System. Int. J. Mol. Sci. 2018, 19, 2504. [Google Scholar] [CrossRef]
- Kajta, M.; Wnuk, A.; Rzemieniec, J.; Lason, W.; Mackowiak, M.; Chwastek, E.; Staniszewska, M.; Nehring, I.; Wojtowicz, A.K. Triclocarban Disrupts the Epigenetic Status of Neuronal Cells and Induces AHR/CAR-Mediated Apoptosis. Mol. Neurobiol. 2018, 56, 3113–3131. [Google Scholar] [CrossRef] [PubMed]
- Stading, R.; Chu, C.; Couroucli, X.; Lingappan, K.; Moorthy, B. Molecular role of cytochrome P4501A enzymes in oxidative stress. Curr. Opin. Toxicol. 2020, 20-21, 77–84. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, J.; Tao, Y.; Ji, C.; Aniagu, S.; Jiang, Y.; Chen, T. AHR/ROS-mediated mitochondria apoptosis contributes to benzo[a]pyrene-induced heart defects and the protective effects of resveratrol. Toxicology 2021, 462, 152965. [Google Scholar] [CrossRef]
- Jenny, M.J.; Karchner, S.I.; Franks, D.G.; Woodin, B.R.; Stegeman, J.J.; Hahn, M.E. Distinct Roles of Two Zebrafish AHR Repressors (AHRRa and AHRRb) in Embryonic Development and Regulating the Response to 2,3,7,8-Tetrachlorodibenzo-p-dioxin. Toxicol. Sci. 2009, 110, 426–441. [Google Scholar] [CrossRef]
Figure 1.
Effects of the motor behavior of zebrafish larvae exposed to PFOSA. (A)Survival rates. (B)Hatching rates. (C) Representative locomotor traces in two light-dark cycles in larval zebrafish. (D) Total distance and average speed in two light-dark cycles. PFOSA 6.25, 12.5, 25, 50: PFOSA at different concentrations (μg/L). * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
Figure 1.
Effects of the motor behavior of zebrafish larvae exposed to PFOSA. (A)Survival rates. (B)Hatching rates. (C) Representative locomotor traces in two light-dark cycles in larval zebrafish. (D) Total distance and average speed in two light-dark cycles. PFOSA 6.25, 12.5, 25, 50: PFOSA at different concentrations (μg/L). * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
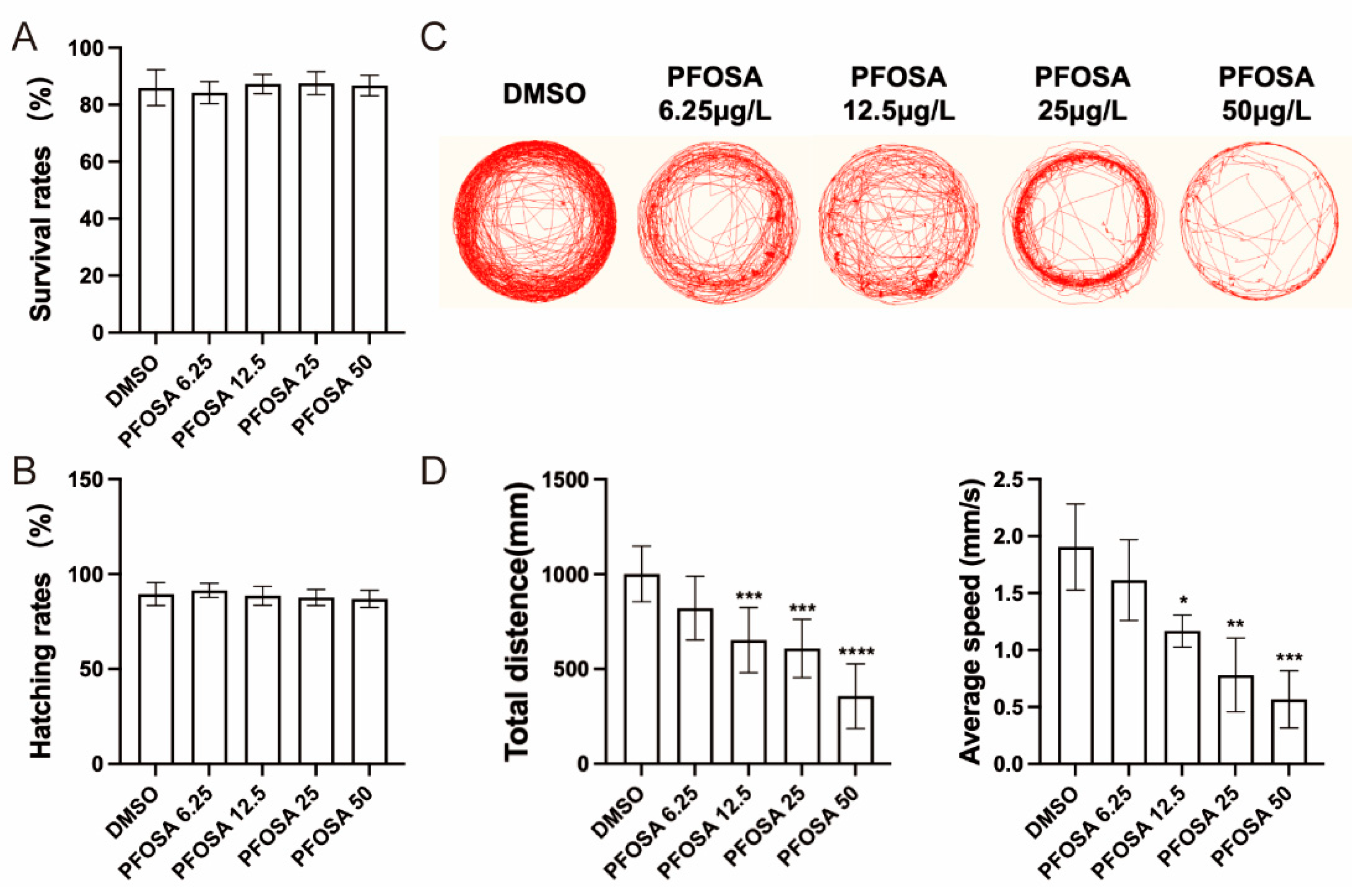
Figure 2.
Effect of zebrafish embryo exposed to PFOSA on neurodevelopment-related genes. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
Figure 2.
Effect of zebrafish embryo exposed to PFOSA on neurodevelopment-related genes. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
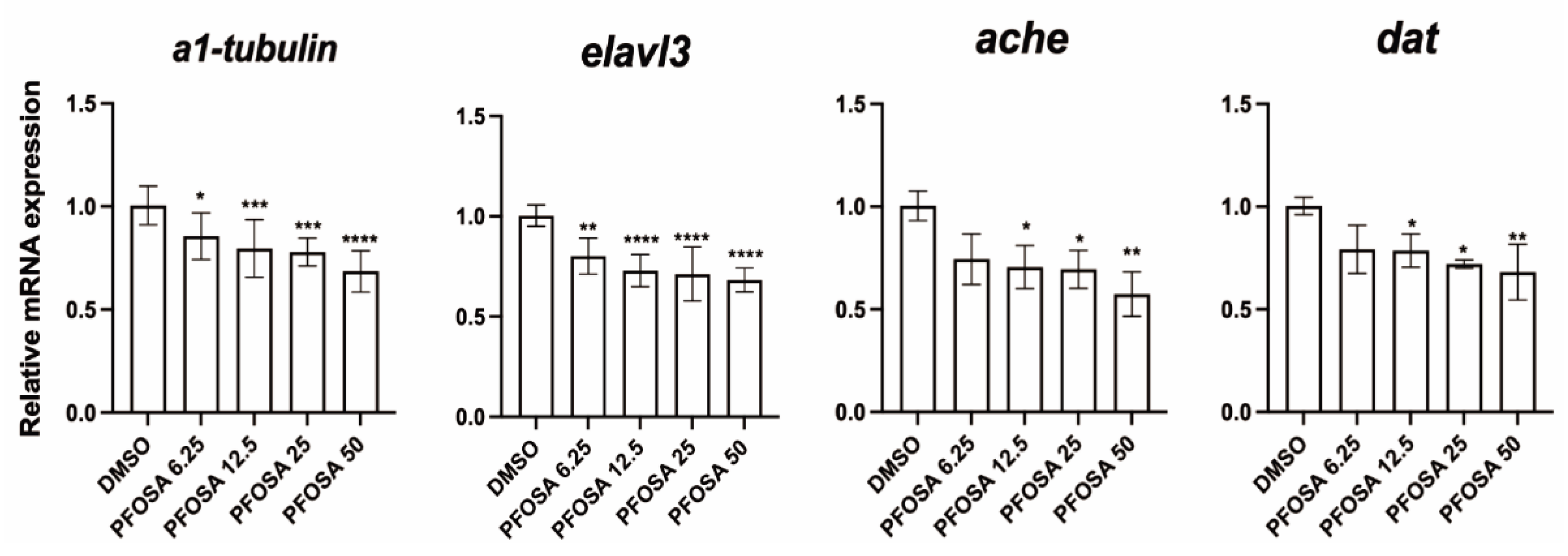
Figure 3.
PFOSA causes oxidative stress and apoptosis in the brain of zebrafish embryos. (A) Intracellular ROS levels. (B) The expression levels of nrf2, sod1, sod2, tp53 and bax in zebrafish embryos from different treatment groups. (C) Apoptosis detected by AO staining. All data are expressed as mean ± standard deviation. *, significant difference compared to the control group (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).
Figure 3.
PFOSA causes oxidative stress and apoptosis in the brain of zebrafish embryos. (A) Intracellular ROS levels. (B) The expression levels of nrf2, sod1, sod2, tp53 and bax in zebrafish embryos from different treatment groups. (C) Apoptosis detected by AO staining. All data are expressed as mean ± standard deviation. *, significant difference compared to the control group (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).
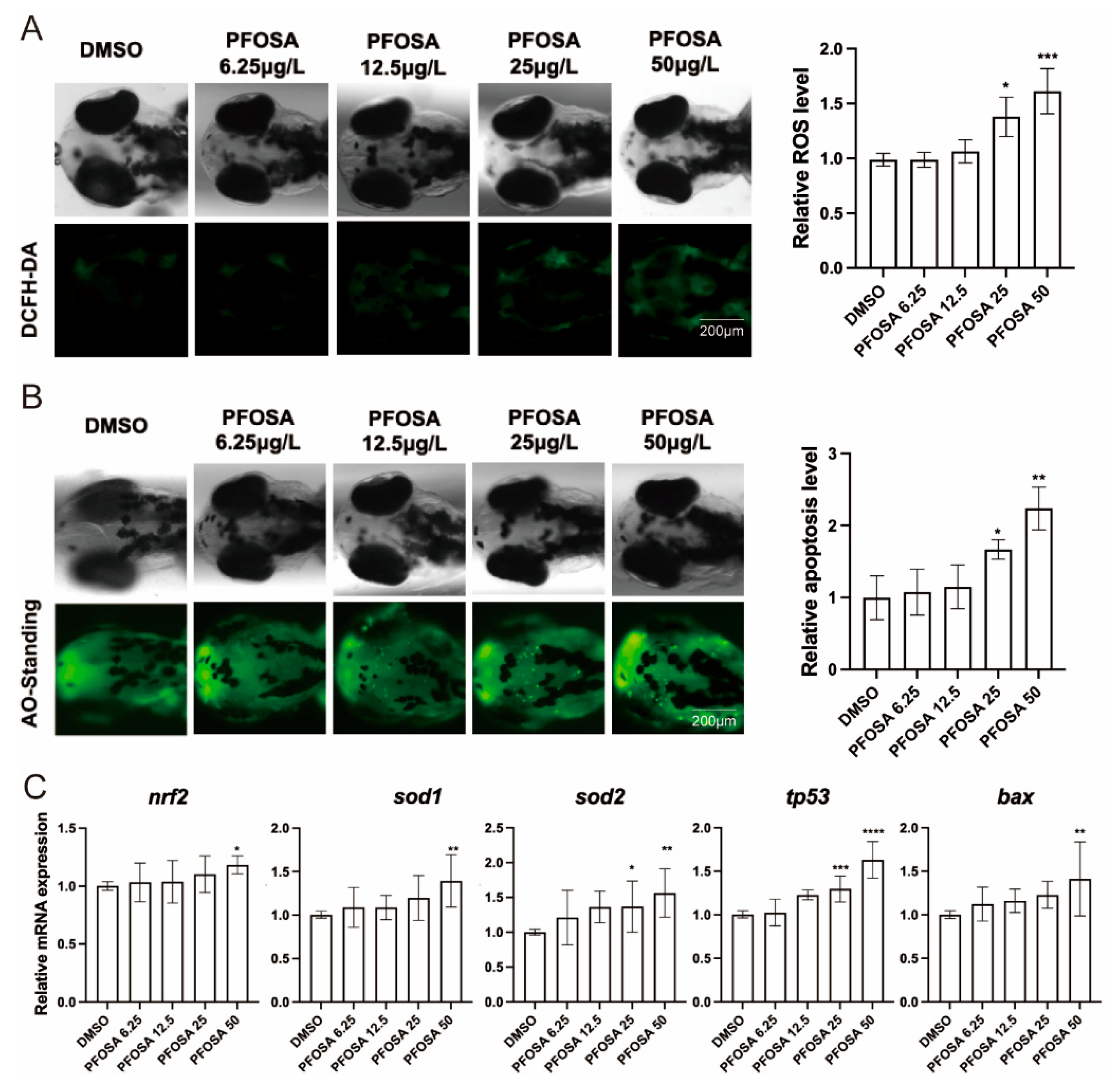
Figure 4.
The AhR/ROS axis mediates PFOSA-induced apoptosis and neurodevelopmental toxicity in zebrafish embryos. (A) Relative mRNA levels of AhR downstream genes. (B) Intracellular ROS levels. (C) Apoptosis detected by AO staining. (D) Representative locomotor traces, total distance, and average speed. (E) Relative mRNA levels of genes involved in neurodevelopment. (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).
Figure 4.
The AhR/ROS axis mediates PFOSA-induced apoptosis and neurodevelopmental toxicity in zebrafish embryos. (A) Relative mRNA levels of AhR downstream genes. (B) Intracellular ROS levels. (C) Apoptosis detected by AO staining. (D) Representative locomotor traces, total distance, and average speed. (E) Relative mRNA levels of genes involved in neurodevelopment. (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).
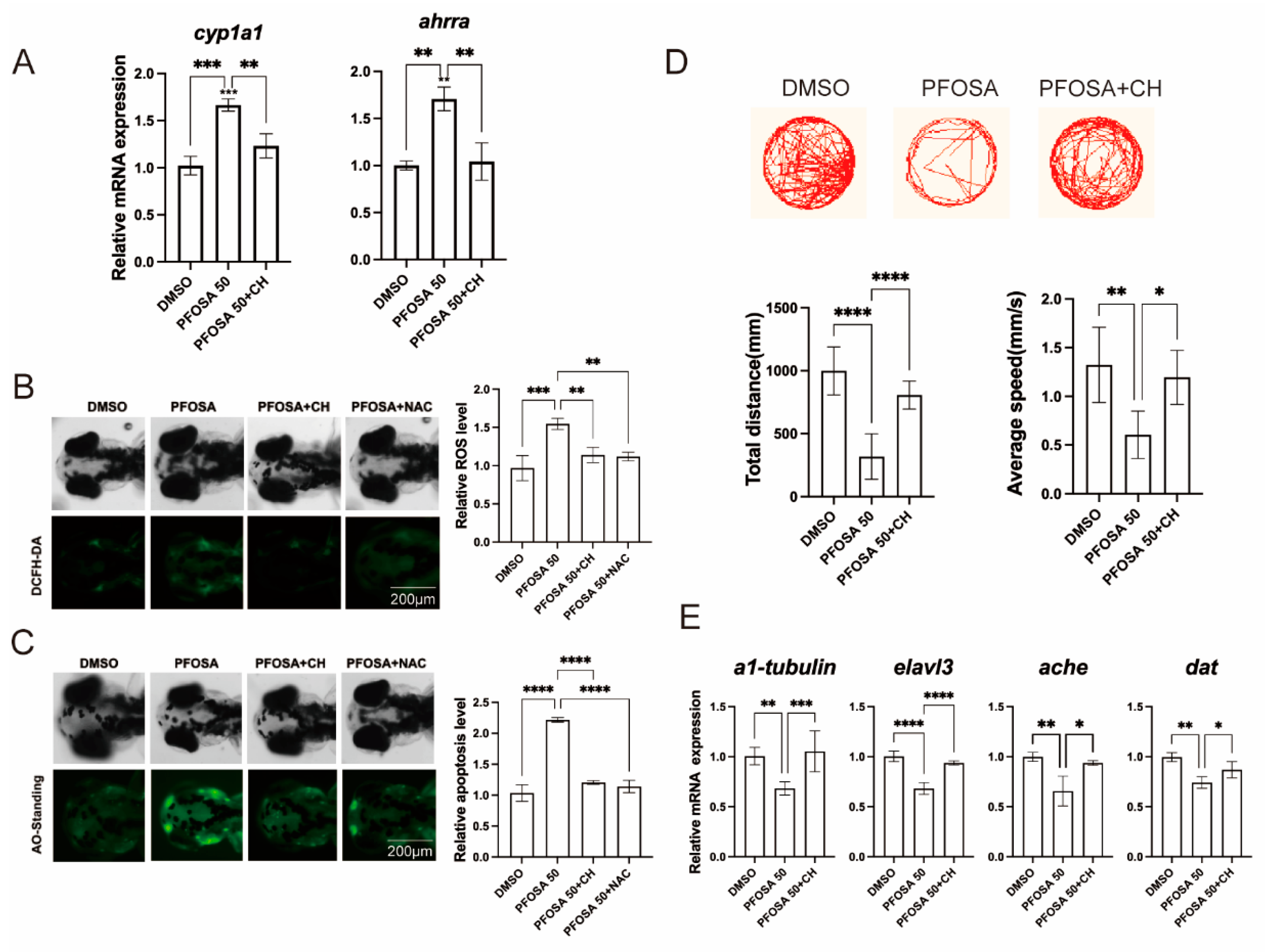

Table 1.
Primer sequences.
| Genes | GenBank No. | Forward (5’-3’) |
| β-actin | NM_131031.2 | CGAGCAGGAGATGGGAACC CAACGGAAACGCTCATTGC |
| elavl3 | NM_131449.1 | TGGTCTGCAGTTTGAGACCGTTGA |
| α1-tubulin | NM_194388.3 | AATCACCAATGCTTGCTTCGAGCC TTCACGTCTTTGGGTACCACGTCA |
| dat | NM_131755.1 | AGACATCTGGGAAGGTGGTG ACCTGAGCATCATACAGGCG |
| ache | NM_131846.3 | CCCTCCAGTGGGTACAAGAA GGGCCTCATCAAAGGTAACA |
| nrf2 | NM_182889.1 | TCGGGTTTGTCCCTAGATG AGGTTTGGAGTGTCCGCTA |
| sod1 | NM_131294.1 | CCGGACTATGTTAAGGCCATCT ACACTCGGTTGCTCTCTTTTCTCT |
| sod2 | NM_199976.1 | GTCGTCTGGCTTGTGGAGTG TGTCAGCGGGCTAGTGCTT |
| tp53 | NM_001271820.1 | CCCGGCGATCATGGATTTAG CCACATGCTCGGACTTCTTATAG |
| bax | NM_131562.2 | GGCTATTTCAACCAGGGTTCC TGCGAATCACCAATGCTGT |
| cyp1a1 | NM_131879.2 | GCATTACGATACGTTCGATAAGGAC GCTCCGAATAGGTCATTGACGAT |
| ahrra | NM_001035265.2 | GCGCATCAAGAGCTTCTGCAGCGTGTT CCACTGACGACCAGCGCAAACCCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



