
Submitted:
10 July 2025
Posted:
11 July 2025
You are already at the latest version
Abstract
Biofilms of uropathogenic bacteria, especially biofilms of Pseudomonas aeruginosa due to the appliance of medical devices such as urinary catheters, cause complications in the treatment of urinary tract infections (UTI). Therefore, the search for efficient control options for biofilms is of great medical interest. This work aims to estimate the use of natural substances as an alternative to antibiotics to mitigate the formation of biofilms also regarding the spreading resistance of uropathogenic bacteria to antibiotics. In this work, due to the complexity of its biofilms, P. aeruginosa ATCC 27853 and P. aeruginosa PA01 were chosen as test organisms. Various natural substances and alkyl glycosides were investigated for their influence on biofilm formation by P. aeruginosa. Biofilm growth on catheter surfaces was determined by the number of colony-forming units per mL(CFU/mL). N-undecyl-α/β-l-fucopyranoside inhibited biofilm formation in P. aeruginosa ATCC 27853, whereas a slight increase in biofilm formation was observed in P. aeruginosa PA01. Significant inhibition of P. aeruginosa biofilm formation was observed with the detergents Soluplus®, Prontosan®, and the fungal metabolite terrein.
Keywords:
alkyl glycosides
; antimicrobial
; biofilm
; catheter
; natural substances
; Pseudomonas aeruginosa
; urinary tract infection
1. Introduction
The biofilms of Pseudomonas aeruginosa pose a serious problem in urological practice. It affects 2 million people every year worldwide, of which 90 thousand die from P. aeruginosa infection [1]. In this work, P. aeruginosa Strain ATCC 27853 serves as test organism, which forms tenacious biofilms [2]. Biofilms in pathogens – such as P. aeruginosa – are a main cause of chronic infections that are difficult to treat properly. This is also the case in the context of urinary tract infections. Urinary catheters are in app. 75% the cause of nosocomial urinary tract infections [3,4,5]. Biofilm formation is a complex process triggered by the nature of the surface, by the interaction of the individual bacterial cells with each other, by Quorum Sensing [3,5,6], and by receptor molecules on biological surfaces [7,8]. Formation of biofilms is a fast process. Within seconds, P. aeruginosa can increase the concentration of the second messenger c-di-GMP. This leads to the activation of the P. aeruginosa- c-di-GMP – receptor FimW followed by the adherence on surfaces and colonization as a biofilm. In addition, asymmetrical cell divisions take place after surface colonization. This leads to the emergence of “spreaders” which disseminate and “strikers” which cause tissue damage [9]. Biofilms formed by P. aeruginosa consist of a matrix of extracellular DNA and polysaccharides. Programmed cell death and autolysis build up and remodel the matrix structures. This is reminiscent of apoptosis, as it occurs in eucaryotes. Leading to polymer-free spaces in the matrix, free P. aeruginosa cells located there can re-swarm and form further biofilms [10]. Biofilms act as a protective shield for the pathogenic organism, so that neither the body's own defenses nor medically used antibiotics could be effective in eradicating the pathogen. Once the biofilm is established, antibiotic treatment cannot eradicate bacteria any more regardless of the susceptibility of biofilm-producing P. aeruginosa to a certain antibiotic [11,12]. As described in the literature, the P. aeruginosa strain PA01 reacts to the presence of aminoglycosides [13] in LB-medium with increased formation of alginate, an exopolysaccharide or with increased biofilm formation. Several strategies exist already against biofilms, such as antibiotic-treated surfaces, for example surfaces secreting silver ions to the medium, or surfaces with anti-adhesive properties [6]. These strategies have, however, not proven clinical relevance, up to date. Where antibacterial, biofilm-preventing strategies do not lead to therapeutic success, biofilm-contaminated surfaces or tissues need to be removed physically. Literature describes micelles and liposomes loaded with antibiotics (both hydrophilic and hydrophobic substances) used against biofilms caused by pathogenic bacteria. Micelles and liposomes already used in medicine have a quite complex structure, albeit with biocompatible components. Some stabilizing polymer components such as polyethylene glycol (PEG) derivatives [3,14,15] are used in micellar systems, however they can cause allergies [3,15]. Another amphiphilic detergent useable for stabilization of micelles or liposomes is Solu-plus® [16] showing a better biocompatibility. Hydrophobic biostatics contain already Soluplus® as a micelle-forming agent [17,18]. Saydam et al. mention Soluplus® as an additive in vaccine mixtures [19]. Rhamnolipids as amphiphilic component in liposomes stimulate the uptake of hydrophobic substances by bacteria, such as P. aeruginosa [20]. The high dosing of classical antibiotics needed to treat biofilms can cause severe adverse events [21,22]. Another issue is the emergence of antimicrobial resistances against antibiotics [23,24,25], where the microenvironment of biofilms creates selective conditions for resistance formation. The concentrations of medically applied antibiotics do not reach the inhibiting/bactericidal level inside a biofilm. In this manner, locally selective concentrations below minimal inhibitory/bactericidal concentrations arise, and resistances can evolve within hours until few days [24]. Due to that, science focuses on plant agents, example given essential oils [26] or polyphenols as curcumin [27] and resveratrol [28]. Polyphenols showed in various research projects unspecific reactions and ambiguous results [29,30], but for curcumin an inhibition-potential of quorum sensing was shown in silico [31]. However in vitro no effect was proven yet [32]. On the other hand, curcumin is recognized as safe by the Food and Drug Administration (FDA) [32]. Under light irradiation with wavelength of 405 nm, curcumin showed an increased inhibition of biofilm by P. aeruginosa. A smaller but still inhibiting effect without the influence of light was also observed [33]. Monolaurin shows antibacterial effects, but only against gram-positive bacteria [34,35]. Substrates for LecB of P. aeruginosa are l-fucose- and d-mannose ligands enabling adhesion to the host tissue surface and consequently biofilm formation by P. aeruginosa. Monovalent low molecular weight derivatives of d-mannose and l-fucose thus being possible inhibitors for biofilm formation of P. aeruginosa [36,37]. The antibacterial agent terrein (4,5-dihydroxy-3-(1-propen-1-yl)-2-cyclopenten-1-one) from Aspergillus terreus acts effectively against the development of P. aeruginosa biofilms. It antagonizes quorum sensing of P. aeruginosa acting as dual inhibitor of QS and c-di-GMP signaling [38]. However, terrein does not affect P. aeruginosa cell growth [38]. The chemical synthesis of terrein is difficult and tedious [39,40,41]. The easiest access to it is the synthesis from l-tartaric acid [42] or isolation from terrein producing A. terreus [43].
The aim of this work was to analyze the capacities of natural substances in prevention of growth and removal of biofilms of uropathogenic bacteria in urinary catheters and thus find novel agents. Our consideration was that detergents can shuttle poorly soluble substances through the outer membrane of the gram-negative P. aeruginosa ATTC 27853. Due to that, the poorly soluble natural substances curcumin, resveratrol, and monolaurin were investigated together with Soluplus® and in the case of monolaurin additionally together with polyhexanide (Prontosan®) for their inhibitory effect against biofilm formation by P. aeruginosa. We have synthesized n-undecyl glycosides of d-mannose and l-fucose according to the modified Fischer synthesis [44] and tested both as substrates for LecB of P. aeruginosa. The fungal metabolite terrein showed the highest efficiency against biofilm formation by P. aeruginosa alone and together with the detergent polyhexanide (Prontosan®).
2. Materials and Methods
2.1. Organisms
Pseudomonas aeruginosa ATCC 27853 was obtained from ATCC (American Type Culture Collection; Manassas, VA, USA), P. aeruginosa PA01 from DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen; Braunschweig, Niedersachsen, Germany).
2.2. Growth Media and Anti-Biofilm Substances
Artificial urine (AU) as medium for P. aeruginosa – test cultures were freshly prepared according to the recipe mentioned in the literature [26,45]. Other media used for cultivation, storage, and testing of P. aeruginosa were glycerol ≥99% (Merck Millipore # CAS 56-81-5), Lysogeny Broth-Medium (LBM) (Serva Electrophoresis GmbH # Cat-No. 48501.01), Tryptone Soy Broth (TSB) (Oxoid # Code: CMO129B). Dulbecco’s phosphate buffered saline (PBS) was purchased from Capricorn Scientific GmbH (# Cat-No: PBS-1A). For chemical syntheses and their monitoring l-(-)-fucose (Carl Roth # CAS No. 2438-80-4), pH-Indicator rod pH-Fix 0-14 (Carl Roth # Art. No. 0549.2), para-n-dodecyl benzenesulfonic acid (Sigma-Aldrich # CAS No. 121-65-3), ethyl acetate (Sigma-Aldrich # CAS-No. 141-78-6), d-(+)-mannose (Sigma-Aldrich # CAS-No. 3458-28-4), n-undecanol (Sigma-Aldrich # CAS-No. 112-42-5), ethanol ≥99,8% (Merck Millipore # CAS 64-17-5), phosphomolybdic acid monohydrate (Merck Millipore # CAS 51429-74-4), sodium hydroxide (Merck Millipore # CAS-No.: 1310-73-2), silicagel 60 (Merck Millipore # CAS No. 7631-86-9), TLC silicagel 60 Aluminum Sheets 5 × 10 cm (Merck Millipore # Cat-No. 1.16835.0001) were used. Piperacillin (Sigma-Aldrich # CAS-No. 66258-76-2) served as a comparison to assess efficacy.
As biofilm inhibiting substances curcumin (Merck Millipore # CAS 458-37-7), monolaurin (TCI # CAS RN®: 142-18-7), resveratrol (TCI # CAS RN®: 501-36-0) were used. Other substances tested as inhibitors were: Soluplus®, a polyvinyl caprolactam–polyvinyl acetate–polyethylene glycol graft copolymer (BASF SE Ludwigshafen am Rhein Germany), polyhexanide (Prontosan®) (B. Braun Melsungen AG, Melsungen, Germany), and terrein (AdipoGen # CAS-No. 582-46-7). For terrein and n-undecyl glycosides, synthesized by us, dimethyl sulfoxide (DMSO) (Sigma-Aldrich # CAS-No. 67-68-5) served as solvent (Table 1).
2.3. Chemical Syntheses
2.3.1. n-Undecyl-α-d-mannopyranoside (MW = 334.35g)
1.04g α/β-d-mannose (5.77 mmol), 11.98 mL n-undecanol (9.94g = 57.72 mmol), 889 μL (942 mg = 2.89 mmol) para-n-dodecylbenzenesulfonic acid and 100 μL fully desalinated H2O were stirred in a closed vessel at 60°C and 700 rpm for 24 h. After the end of the reaction, the initially white, cloudy mixture becomes clear. To monitor the reaction process thin layer chromatography was performed (eluent ethyl acetate/ethanol 70:30). Unreacted n-undecanol was removed via column chromatography on silica gel eluting with ethyl acetate/ ethanol (70:30). To remove para-n-dodecylbenzenesulfonic acid residues, the crude n-undecyl-α-d-mannopyranoside was dissolved in 10 mL of ethanol at 55°C and after cooling, addition of ice-cold ethyl acetate led to crystallization of the desired product. The resulting crystal slurry was filtered off, washed with ice-cold ethyl acetate, and the product was dried in the fume hood [44]. The product is a white crystalline solid. Yield: 35% related to α/β-d-mannose used, see 3.1.1. The identity of the product was confirmed by mass spectrometry and nuclear magnetic resonance spectroscopy (NMR). 1H NMR-analysis showed diastereomeric purity of for the α-anomer.
2.3.2. n-Undecyl-α/β-l-fucopyranoside (MW = 318.35 g)
1.96 g α/β-l-fucose (11.95 mmol), 24.69 mL n-undecanol (20.49 g = 118.90 mmol), 1.832 mL (1,942 g = 5.98 mmol) para-n-dodecylbenzenesulfonic acid and 190 μL fully desalinated H2O were stirred in a closed vessel at 60°C and 700 rpm for 24 h. The initial yellowish white suspension was a slightly cloudy mixture after the end of the reaction. To monitor the reaction process thin layer chromatography was performed (eluent ethyl acetate/ethanol 90:10). Unreacted n-undecanol was removed via column chromatography on silica gel eluting with ethyl acetate/ethanol (100:0 –> 90:10). [44]. The product is a white crystalline solid. Yield: 60% related to α/β-l-fucose used, see 3.1.2. The identity of the product was confirmed by mass spectrometry and nuclear magnetic resonance spectroscopy (NMR). 1H NMR-analysis showed an anomeric ratio of ≈ 2:1 in favor of the α -anomer.
2.4. Bacterial Incubation
For incubating bacterial cultures an IncuLine IL 112 Prime.390-0910 (VWR International GmbH Darmstadt Germany) and a Shaking Incubator IKA® KS 4000 i Control (IKA-Werke GmbH & Co. KG Staufen Germany) were used. To monitor bacterial growth a Laxco MicroSpek™ DSM-Series Cell Density Meter (LAXCO Bothell, WA 98012 USA) and semi-micro-Cuvettes Polystyrene 45 * 12 mm (Sarstedt AG & Co.KG Nümbrecht Germany) were used.
2.5. Determination of Minimal Inhibitory Concentration
Determination of minimal inhibitory concentration (MIC)-Values was performed with 96-Well-Plates Checkerboard TC-Plate 96 Well flat bottom Standard (Sarstedt AG & Co.KG Nümbrecht Germany) and a MultiScanGO (Thermo Fisher Scientific GmbH Langerwehe Germany). MIC-values were determined according to EUCAST standards [46,47] of antibacterial agents by broth dilution. From the agent to be tested a two-fold dilution series with a volume of 160 µL per well is made in AU. Then every well containing agent was inoculated with 10 µL of PBS-buffer diluted P. aeruginosa ATCC 27853 -suspension (1:200 v/v). As positive control served medium without agent inoculated, negative control served medium without bacteria and agent. After incubation overnight at 37°C turbidity was measured at λ = 600 nm. MIC means the lowest concentration where no additional turbidity (corresponding to bacterial growth) compared to positive control can be detected.
2.6. Urinary Catheters and Processing
For catheter testing Rüsch-Gold- Latex-Catheters (Teleflex Medical GmbH Fellbach Germany) served as substrate for biofilm growth. An Ultrasonic device Bandelin SONOREX TK30 50 kHz (BANDELIN electronic GmbH & Co. KG Berlin Germany) and a Vortex Scientific Industries Vortex-Genie 2 G560 S/N 2-417489 (Thermo Fisher Scientific GmbH Langerwehe Germany) were used to process the biofilms and subsequently determine the CFU count (see 2.8.).
2.7. Catheter Biofilm Experiments
Preparation of P. aeruginosa ATCC 27853 for catheter experiment: From the – 80°C glycerol stock of P. aeruginosa ATCC 27853 a small inoculum was streaked on LB agar and incubated for 16 h at + 37°C. Subsequently, a single colony of the agar plate culture was incubated in 5 mL AU in a 100 mL Erlenmeyer flask for 16 h at 37°C and at 180 rpm in the shaker incubator as primary culture (Figure 1a – 1c).
2.7.1. General Procedure Catheter Experiment
The primary culture of P. aeruginosa ATCC 27853 was diluted with sterile PBS-buffer to an optical density (ODλ=600 nm) = 0.02. To 8 mL of medium and three 1 cm long catheter segments as technical replicates 100 µL of the PBS-buffer-diluted P. aeruginosa ATCC 27853 suspension was added. Before adding P. aeruginosa ATCC 27853, the tested substance was added in appropriate concentration into the medium. Thereafter incubation over 24 h to 36 h at 37°C was performed and after processing of grown biofilms CFU were counted (see 2.8). Each test series with a specific substance or combination of substances was biologically replicated three times, (Figure 1c – 1f).
2.7.2. Influence of the Culture Medium
Due to results shown for Proteus mirabilis [48], differences in growth behavior of P. aeruginosa ATCC 27853 in AU and LB-Medium were estimated. To 8 mL of AU and LB medium without additives and to three 1-cm-long latex catheter segments respectively, P. aeruginosa ATCC 27853 -suspension with an ODλ=600nm = 0.02 in PBS-buffer was added. After incubation for 24 h at 37 °C colony counting follows, (Figure 2a).
2.7.3. Curcumin, Monolaurin, Resveratrol, Each in Combination with Soluplus
To 8 mL AU and three 1 cm long catheter segments 512 µg/mL curcumin together with 5120 µg/mL Soluplus were added and incubated over 24 h at 37°C in the presence of P. aeruginosa ATCC 27853 according to the general procedure. As controls, three 1 cm long catheter segments and P. aeruginosa ATCC 27853 were incubated without additive in AU and with 5120 µg/mL Soluplus without further additive (Figure 2b). In the same manner 512 µg/mL AU resveratrol together with 5120 µg/mL AU Soluplus (Figure 2c) and 512 µg/mL AU monolaurin together with 48 µg/ml AU Prontosan (Figure 2d) and together with 128 µg to 5120 µg/mL AU Soluplus (Figure 2e) are tested. Controls with curcumin, resveratrol, and were not feasible due to their insolubility in AU.
2.7.4. n-Undecylglycosides
n-Undecyl-α-d-mannopyranoside was tested against biofilm forming by P. aeruginosa ATCC 27853 in concentrations of 32 µg/mL AU to 512 µg/mL AU following the general procedure (Figure 3a). n-Undecyl-α/β-l-fucopyranoside was tested in concentrations of 32 µg/mL AU to 512 µg/mL AU following the general procedure, against biofilm forming by P. aeruginosa ATCC 27853 (Figure 3b) and by P. aeruginosa PA01 (Figure 3c).
2.7.5. Terrein
Terrein, an antimicrobial metabolite from the mould Aspergillus terreus, was tested in concentrations of 32 µg/mL AU to 512 µg/mL AU following the general procedure (Figure 5a).
2.7.6. Co-Effects of Terrein and Polyhexanide
To estimate possible synergism, 256 µg/mL AU terrein together with 64 µg/mL AU polyhexanide were tested on mitigation of biofilm formation by P. aeruginosa ATCC 27853. Because keratinocytes tolerate polyhexanide well in concentrations up to 175 µg/mL [49] we have chosen polyhexanide as co-agent instead of Soluplus®, (Figure 5b).
2.8. Estimation of Colony Forming Units
The 1 cm long biofilm-covered catheter was washed 3 times with sterile 1000 μL PBS buffer. Then the application of 50 kHz ultrasound for 5 min to the catheter-piece, vortex-shaking for 2 min, 50 kHz ultrasound for 5 min again and vortex-shaking for 1 min followed the washing. After discarding of the catheter piece, the obtained suspension was diluted in series of 10. Of suitable concentrations (10-4 to 10-7), 100 μL of bacterial suspension was spread on LB agar plates and incubated "overnight" at + 37°C. Then the colonies were counted [49,50]. It must be considered that only one tenth of a mL is spread on agar.
2.9. Statistical Analysis
For statistical analyses served GraphPad Prism 10, version: GraphPad Prism 10.4.2.. Statistical significance was set as p ≤ 0.05, with α = 0.05. The degree of significance is shown using * for p ≤ 0.05, ** p ≤ 0.01, ***p ≤ 0.001 and ****p ≤ 0.0001. For statistical comparison of two data sets with GraphPad Prism 10.4.2 a 2-tailed t-test was used. One way ANOVA test served for analysis of more than three data sets. ANOVA-Dunnet test served to compare multiple data sets with a single control. The Tukey test was used to compare every data set with every other.
3. Results
3.1. Chemical Syntheses
3.1.1. n-Undecyl-α-d-Mannopyranoside
The yield for anomerically pure n-undecyl-α-d-mannopyranoside was 0.68 g (2.02 mmol; 35%) related to 1.04 g (5.77 mmol) α/β-d-mannose used in the synthesis. HRMS (ESI): m/z calculated for C17H34O6Na [M+Na]+: 357.2248; found: 357.2247. 1H NMR (600 MHz, CD3OD): δ 4.73 (d, J = 1.8 Hz, 1H, H-1), 3.82 (dd, J = 11.8, 2.4 Hz, 1H, H-6a), 3.78 (dd, J =3.4, 1.7 Hz, 1H, H-2), 3.75 – 3.66 (m, 3H, H-3/–OCH2CH2/H-6b), 3.61 (app t, J = 9.6 Hz, 1H, H-4), 3.52 (ddd, J = 9.8, 5.8, 2.4 Hz, 1H, H-5), 3.41 (dt, J = 9.6, 6.3 Hz, 1H, –OCH2CH2), 1.64 – 1.53 (m, 2H, –OCH2CH2), 1.43 – 1.20 (m, 16H, 8 CH2), 0.90 (t, J = 6.9 Hz, 3H, CH3) ppm. 13C{1H} NMR (150 MHz, CD3OD): δ 101.5 (C-1), 74.5 (C-5), 72.7 (C-3), 72.3 (C-2), 68.6 (C-4), 68.6 (–OCH2), 62.9 (C-6), 33.1, 30.8, 30.7, 30.6, 30.6, 30.5, 27.4, 23.7, (9 CH2) 14.4 (CH3) ppm (supplementary data).
3.1.2. n-Undecyl-α/β-l-Fucopyranoside
The yield for n-undecyl-α/β-l-fucopyranoside was 2.297 g (7.213 mmol; 60%) related to the 1.96 g (11.95 mmol) of α/β-l-fucose used, α/β-ratio was ≈ 2:1 according to 1H NMR analysis. HRMS (ESI): m/z calculated for C17H34O5Na [M+Na]+: 341.2299; found: 341.2297. 1H NMR (600 MHz, CD3OD): δ 4.73 (d, J = 2.7 Hz, 1H, H-1α), 4.17 (d, J = 7.5 Hz, 0.5H, H-1β), 3.97 – 3.91 (m, 1H, H-5α), 3.83 (dt, J = 9.5, 6.8 Hz, 0.5H, β–OCH2CH2), 3.74 – 3.70 (m, 2H, H-2α/H-3α), 3.68 – 3.58 (m, 3H, α–OCH2CH2/H-4α/H-6β), 3.51 (dt, J = 9.5, 6.7 Hz, 0.5H, β–OCH2CH2), 3.47 – 3.41 (m, 2H, α–OCH2CH2, H-4β, H-4β), 1.72 – 1.50 (m, 3H, α–OCH2CH2/β–OCH2CH2), 1.43 – 1.23 (m, 25.5H, α-CH2/β-CH2/H-6β), 1.21 (d, J = 6.6 Hz, 3H, H-6α), 0.90 (t, J = 6.9 Hz, 4.5H, α-CH3/β-CH3) ppm. 13C{1H} NMR (150 MHz, CD3OD): δ 104.8 (C-1β), 100.5 (C-1α), 75.2 (C-5β), 73.6 (C-4α), 73.1 (C-3β), 72.3 (C-2β), 71.8 (C-4β), 71.7 (C-3α), 70.8 (β–OCH2), 70.0 (C-2α), 69.3 (α–OCH2), 67.5 (C-5α), 33.1, 30.9, 30.8, 30.7, 30.6, 30.6, 30.6, 30.5, 27.3, 27.1, 23.7 (α-CH2/β-CH2), 16.8 (C-6β), 16.7 (C-6α), 14.4 (α-CH3/β-CH3) ppm (supplementary data).
3.2. MIC-Values
MIC-Values are shown in Table 2. For Piperacillin the MIC Value of 2 – 4 µg/mL in iron-depleted Mueller-Hinton broth according to EUCAST -standards was confirmed with our result of 4 µg/mL AU as MIC for P. aeruginosa ATCC 27853. In combination with Soluplus® (polyvinyl caprolactam–polyvinyl acetate–polyethylene glycol graft copolymer) the MIC for Piperacillin did not change. Surprisingly the detergent Soluplus® did not show any bacteria inhibiting effect. Polyhexanide (Prontosan®) showed an inhibitory effect against P. aeruginosa cells. The determined MIC value was 16/32 µg/mL AU and corresponds to the value reported in the literature. For the MIC value of terrein, no comparative value could be found in the literature. When terrein and Prontosan were combined, a reduction in the MIC was observed for terrein. Monolaurin showed no bacteria inhibiting effect. The combination with Soluplus® also did not cause any bacteriostatic effect of monolaurin. The same was observed when combining curcumin or resveratrol with Soluplus®. Since no bacteria inhibiting effect was expected for n-undecyl-α-d-mannopyranoside, n-undecyl-α/β-l-fucopyranoside, and curcumin, which is insoluble in aqueous media, no MIC determinations were carried out for these four substances.
3.3. Dependence of Biofilm Formation on the Culture Medium
Primary culture of P. aeruginosa ATCC 27853 cultivated in AU and LBM showed in both cases biofilm formation. There was no significant difference in the colony numbers obtained from these biofilms. (Figure 2a).
3.4. Growth Inhibition of Pseudomonas aeruginosa-Biofilms
3.4.1. Curcumin, Monolaurin, Resveratrol Each with Detergent
Curcumin in combination with Soluplus® caused no change in the inhibitory effect of Soluplus against biofilm forming by P. aeruginosa ATCC 27853, although Soluplus alone already showed a strong inhibitory effect, this was not further enhanced by curcumin (Figure 2b). Addition of resveratrol also did not significantly increase the strong Soluplus effect alone on biofilm formation by P. aeruginosa ATCC 27853, (Figure 2c). Further tests with curcumin and resveratrol have therefore not been carried out. Monolaurin showed in combination with Prontosan® no additional effect, (Figure 2d), with Soluplus® additive inhibition against biofilm formation by P. aeruginosa ATCC 27853 proved to be non-significant, (Figure 2e).
3.4.2. n-Undecyl-α-d-mannopyranoside
n-Undecyl-α-d-mannopyranoside did not inhibit biofilm formation by P. aeruginosa ATCC 27853. At concentration of 256 µg/mL AU n-undecyl-α-d-mannopyranoside caused a significant increase in biofilm formation of 80%, (Figure 3a).
3.4.3. n-Undecyl-α/β-l-fucopyranoside
n-Undecyl-α/β-l-fucopyranoside showed 58.3% inhibition as an inhibitor of biofilm formation by P. aeruginosa ATCC 27853 at concentration of 512 µg/mL, (Figure 3b). In contrast, n-undecyl-α/β-l-fucopyranoside did not inhibit biofilm formation of P. aeruginosa PA01, and at a concentration of 256 µg/mL it caused an increase in biofilm formation. Between n-undecyl-α/β-l-fucopyranoside -concentrations of 64 µg/mL to 256 µg/mL, an increase in biofilm formation of 87% occurred, (Figure 3c).
3.4.4. Terrein
Terrein showed from a concentration of 256 mg/mL 55.6% significant inhibition of biofilm formation by P. aeruginosa ATCC 27853, and at 512 µg/mL AU terrein caused a nearly complete inhibition of biofilm formation, (Figure 4a).
3.4.6. Co-Effects of Terrein and Polyhexanide
Together terrein and the disinfectant Polyhexanide (Prontosan®) tested against P. aeruginosa ATCC 27853 -biofilm formation seem to have a synergistic effect against biofilm formation by P. aeruginosa ATCC 27853, but significance is not confirmable statistically, (Figure 4b).
4. Discussion
We investigated the effect of various natural products regarding their inhibitory effect on the biofilm formation of Pseudomonas aeruginosa ATCC 27853. The influence of the two media LBM and AU on the growth of P. aeruginosa ATCC 27853 was not significant. We have shown that the polyphenols curcumin and resveratrol showed a significant effect against biofilm formation by P. aeruginosa ATCC 27853 in the presence of detergent. The working group of A. U. Khan has shown that curcumin has an inhibitory effect against biofilm formation of P. aeruginosa MTCC 3541. This effect was significant without light irradiation and additionally intensified by light irradiation [35]. Curcumin, resveratrol, and monolaurin are nearly insoluble in the culture media used. Therefore, inhibition tests with curcumin, resveratrol, or monolaurin alone were not feasible. The effectiveness in combination with the cleaning and disinfectant Soluplus® cannot be evaluated based on the results obtained so far. The reason for this is that Soluplus® in the concentration necessary to dissolve curcumin override any possible curcumin effect. So, criticism on the use of curcumin against the biofilm formation of (uro-) pathogenic bacteria [29] was confirmed in our studies on P. aeruginosa ATCC 27853. It should be emphasized that further studies with curcumin for targeted medical applications - beyond the certainly justified use in folk medicine as an active ingredient in turmeric roots [35] - have little chance of success, unless a simple method for permanent solution in aqueous media can be established to demask effects, superimposed by the detergent Soluplus® used so far. We observed similar results when using resveratrol. The idea that monolaurin could show an additive effect in combination with Soluplus® enabling monolaurin to cross the outer membrane from gram negative P. aeruginosa ATCC 27853 could not be confirmed. The effect of monolaurin in combination with Soluplus® to inhibit biofilm formation by P. aeruginosa ATCC 27853 was not statistically significant. Together with 48 µg/mL Prontosan, 256 µg/mL monolaurin did not increase effectiveness compared to Prontosan alone. Tested as possible antilectins, n-undecyl-α-d-mannopyranoside showed no inhibitory effect on biofilm formation, but n-undecyl-α/β-l-fucopyranoside showed significant medium effect to attenuate biofilm formation of P. aeruginosa ATCC 27853. On the other hand, n-undecyl-α/β-l-fucopyranoside proved to be not effective against P. aeruginosa PA01. One reason for these results may be the fact that P. aeruginosa has a high genomic diversity among different isolates [51]. Among natural substances tested, terrein considered nontoxic [52] and therefore conceivable as a potential therapeutic agent, however, showed a significant inhibitory effect against biofilm formation by P. aeruginosa ATCC 27853. Moreover polyhexanide (Prontosan®) significantly increased the effect of terrein. It is possible that Prontosan® facilitates the passage of terrein through the outer membrane of P. aeruginosa ATCC 27853. The detergents, for example, may be applied locally in the urinary catheter balloon, as a slow-release device [49]. Terrein, in addition to its inhibitory effect on P. aeruginosa biofilm formation, has been shown to inhibit the migration of breast cancer cells and proved to be non-toxic in the in vitro experiments [53]. However, toxicity studies with bladder epithelial cells are needed to assess possible adverse effects on healthy eukaryotic cells, if clinical application in the urinary tract is envisioned.
5. Conclusions
The screening of natural substances on effectiveness against formation of biofilm is a promising approach to circumvent the development of resistances and adverse effects emerging with antibiotics used. However, some authors are critical on the chances of success in research with apparently multifactorial natural substances. Testing curcumin, resveratrol, and monolaurin without further additives is not feasible, due to problems to dissolve these substances in aqueous media completely. Both biocompatible detergents Soluplus® and polyhexanide (Prontosan®) used by us to circumvent this issue, mask in the necessarily used concentrations possible anti-bacterial effects in case of curcumin, resveratrol, and monolaurin. This may be due to the strong effectiveness of the cleaning agents themselves or to the weak effect of these three substances against P. aeruginosa biofilm formation. Therefore, further efforts to circumvent the main problem of their lack of solubility more efficiently in aqueous media should be performed. Subsequently, further tests can be carried out. The possible antilectin n-undecyl-α/β-l-fucopyranoside is yet to be tested with polyhexanide (Prontosan®) to investigate possible enhancement of effectiveness against biofilm formation of P. aeruginosa ATCC 27853. Another approach is to test the chemically pure anomers of n-undecyl-α/β-l-fucopyranoside against biofilm formation by P. aeruginosa. Due to its strong effectiveness against biofilm formation the fungal metabolite terrein should also be tested against established biofilms, with and without polyhexanide and Soluplus. A further interest is to investigate the effect of terrein against biofilm formation by P. aeruginosa PA01 on catheter surfaces. Another question is to what extent a possible synergism of terrein and n-undecyl-α/β-l-fucopyranoside could mitigate the formation of biofilms. The significant effect that terrein has shown in inhibiting biofilms on latex catheter material is reason enough to investigate other bacteriostatic or bactericidal agents for biofilm inhibition on latex and other catheter materials.
Supplementary Materials
All figures are included in this published manuscript.
Author Contributions
Conceptualization C.V.; data curation C.V. and A.A.; methodology, C.V., A.A., and R.C.W.; project administration, C.V.; formal analysis, C.V., A.A., R.C.W., U.S., F.W.; investigation, C.V., A.A., R.C.W., F.W.; supervision C.V., U.S., F.W.; validation C.V.; visualization, C.V., A.A. and F.W.; resources, R.C.W., F.W. and U.S.; writing-original draft preparation C.V.; writing-review and editing, C.V. A.A., R.C.W., U.S. and F.W. All authors have read and agreed to the published version of the manuscript.
Funding
These investigations received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated and analyzed during this work are included in this published manuscript.
Acknowledgments
We thank Tanja Bloch and Kerstin Wilhelm for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, Xuejie; Gu, Nixuan; Huang, Teng Yi; Zhong, Feifeng; Peng, Gongyong: Pseudomonas aeruginosa: A typical biofilm forming pathogen and an emerging but underestimated pathogen in food processing. Front Microbiol. 2023 Jan 25:13:1114199. [CrossRef]
- Cao, Huiluo; Lai, Yong; Bougouffa, Salim; Xu, Zeling; Yan, Aixin: Comparative genome and transcriptome analysis reveals distinctive surface characteristics and unique physiological potentials of Pseudomonas aeruginosa ATCC 27853. BMC Genomics. 2017 Jun 12;18(1):459. [CrossRef]
- Singha, Priyadarshini; Locklin, Jason; Handa, Hitesh: A review of the recent advances in antimicrobial coatings for urinary catheters. Acta Biomater. 2017 Mar 1:50:20-40. [CrossRef]
- Kranz, Jennifer; Schmidt, Stefanie; Wagenlehner, Florian; Schneidewind, Laila: Catheter-Associated Urinary Tract Infections in Adult Patients. Dtsch Arztebl Int. 2020 Feb 7;117(6):83-88. [CrossRef]
- Zhu, Zhiling; Wang, Ziping; Li, Siheng; Yuan, Xun: Antimicrobial strategies for urinary catheters. J Biomed Mater Res A. 2019 Feb;107(2):445-467. [CrossRef]
- Rasamiravaka, Tsiry; Labtani, Quentin; Duez, Pierre; El Jaziri, Mondher: The formation of biofilms by Pseudomonas aeruginosa: a review of the natural and synthetic compounds interfering with control mechanisms. Biomed Res Int. 2015:2015:759348. [CrossRef]
- Öztürk, Fırat Yavuz; Darcan, Cihan; Kariptaş, Ergin: The Determination, Monitoring, Molecular Mechanisms and Formation of Biofilm in E. coli. Braz J Microbiol. 2023 Mar;54(1):259-277. [CrossRef]
- Sauer, Karin; Camper, Anne K.; Ehrlich, Garth D.; Costerton, J. William; Davies, David G.: Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J Bacteriol. 2002 Feb;184(4):1140-54. [CrossRef]
- Laventie, Benoît-Joseph; Sangermani, Matteo; Estermann, Fabienne; Manfredi, Pablo; Planes, Rémi; Hug, Isabelle et al.: A Surface-Induced Asymmetric Program Promotes Tissue Colonization by Pseudomonas aeruginosa. Cell Host Microbe. 2019 Jan 9;25(1):140-152.e6. [CrossRef]
- Ma, Luyan; Conover, Matthew; Lu, Haiping; Parsek, Matthew R.; Bayles, Kenneth; Wozniak, Daniel J.: Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathog. 2009 Mar;5(3):e1000354. [CrossRef]
- Bagge, Niels; Hentzer, Morten; Andersen, Jens Bo; Ciofu, Oana; Givskov, Michael; Høiby, Niels: Dynamics and spatial distribution of beta-lactamase expression in Pseudomonas aeruginosa biofilms. Antimicrob Agents Chemother. 2004 Apr;48(4):1168-74. [CrossRef]
- Bjarnsholt, Thomas: The role of bacterial biofilms in chronic infections. APMIS Suppl. 2013 May:(136):1-51. [CrossRef]
- Hoffman, Lucas R.; D'Argenio, David A.; MacCoss, Michael J.; Zhang, Zhaoying; Jones, Roger A.; Miller, Samuel I.: Aminoglycoside antibiotics induce bacterial biofilm formation. Nature. 2005 Aug 25;436(7054):1171-5. [CrossRef]
- Coimbra, Maria; Isacchi, Benedetta; van Bloois, Louis; Torano, Javier Sastre; Ket, Aldo; Wu, Xiaojie et al.: Improving solubility and chemical stability of natural compounds for medicinal use by incorporation into liposomes. Int J Pharm. 2011 Sep 20;416(2):433-42. [CrossRef]
- Cox, Fionnuala; Khalib, Khairin; Conlon, Niall: PEG That Reaction: A Case Series of Allergy to Polyethylene Glycol. J Clin Pharmacol. 2021 Jun;61(6):832-835. [CrossRef]
- Hendrik Hardung, Dejan Djuric, and Shaukat Ali: Combining HME & Solubilization: Soluplus® - The Solid Solution Drug Delivery Technology 2010 Vol 10 No 3, 20-27 https://www.researchgate.net/publication/279893959_Combining_HME_solubilization_SoluplusR_-_The_solid_solution.
- Ji, Suping; Lin, Xiao; Yu, Enjiang; Dian, Chengyang; Yan, Xiong; Li, Liangyao et al.: Curcumin-Loaded Mixed Micelles: Preparation, Characterization, and In Vitro Antitumor Activity. J. Nanotechnology 2018, 2018(1):9103120. [CrossRef]
- Shamma, Rehab N.; Basha, Mona: Soluplus®: A novel polymeric solubilizer for optimization of Carvedilol solid dispersions: Formulation design and effect of method of preparation. Powder Technol. 237 2013: 406–414. [CrossRef]
- Saydam, Manolya; Cheng, Woei Ping; Palmer, Nathan; Tierney, Robert; Francis, Robert; MacLellan-Gibson, Kirsty et al.: Nano-sized Soluplus® polymeric micelles enhance the induction of tetanus toxin neutralizing antibody response following transcutaneous immunization with tetanus toxoid. Vaccine. 2017 Apr.25;35(18): 2489–2495. [CrossRef]
- Noordman, Wouter H.; Janssen, Dick B.: Rhamnolipid stimulates uptake of hydrophobic compounds by Pseudomonas aeruginosa. Appl Environ Microbiol. 2002 Sep;68(9):4502-8. [CrossRef]
- Baik, Seo; Lau, Jason; Huser, Vojtech; McDonald, Clement J.: Association between tendon ruptures and use of fluoroquinolone, and other oral antibiotics: a 10-year retrospective study of 1 million US senior Medicare beneficiaries. BMJ Open. 2020 Dec 21;10(12):e034844. [CrossRef]
- Jiang, Meiyan; Li, Hongzhe; Johnson, Anastasiya; Karasawa, Takatoshi; Zhang, Yuan; Meier, William B. et al.: Inflammation up-regulates cochlear expression of TRPV1 to potentiate drug-induced hearing loss. Sci Adv. 2019 Jul 17;5(7):eaaw1836. [CrossRef]
- Boucher, Helen W.; Talbot, George H.; Bradley, John S.; Edwards, John E.; Gilbert, David; Rice, Louis B. et al.: Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America. Clin Infect Dis. 2009 Jan 1;48(1):1-12. [CrossRef]
- Sandegren, Linus: Selection of antibiotic resistance at very low antibiotic concentrations. Ups J Med Sci. 2014 May 19;119(2):103–107. [CrossRef]
- Shlaes, David M.; Sahm, Dan; Opiela, Carol; Spellberg, Brad: The FDA reboot of antibiotic development. Antimicrob Agents Chemother. 2013 Oct;57(10):4605-7. [CrossRef]
- Loose, Maria; Pilger, Emmelie; Wagenlehner, Florian: Anti-Bacterial Effects of Essential Oils against Uropathogenic Bacteria. Antibiotics (Basel). 2020 Jun 25;9(6):358. [CrossRef]
- Lin, Derong; Xiao, Lijuan; Qin, Wen; Loy, Douglas A.; Wu, Zhijun; Chen, Hong; Zhang, Qing: Preparation, characterization and antioxidant properties of curcumin encapsulated chitosan/lignosulfonate micelles. Carbohydr Polym. 2022 Apr 1:281:119080. [CrossRef]
- Qi, Lin; Liang, Rongxin; Duan, Jingjing; Song, Songze; Pan, Yunjun; Liu, Hui et al.: Synergistic antibacterial and anti-biofilm activities of resveratrol and polymyxin B against multidrug-resistant Pseudomonas aeruginosa. J Antibiot (Tokyo). 2022 Oct;75(10):567-575. [CrossRef]
- Aldrich, Courtney; Bertozzi, Carolyn; Georg, Gunda I.; Kiessling, Laura; Lindsley, Craig; Liotta, Dennis et al.: The Ecstasy and Agony of Assay Interference Compounds. J Med Chem. 2017 Mar 23;60(6):2165-2168. [CrossRef]
- Baell, Jonathan; Walters, Michael A.: Chemistry: Chemical con artists foil drug discovery. Nature. 2014 Sep 25;513(7519):481-3. [CrossRef]
- Ding, T., Li, T., Wang, Z., Li, J.: Curcumin liposomes interfere with quorum sensing system of Aeromonas sobria and in silico analysis. Sci Rep. 2017 Aug 17;7(1):8612. [CrossRef]
- Fernandes, Susana; Borges, Anabela; Gomes, Inês B.; Sousa, Sérgio F.; Simões, Manuel: Curcumin and 10-undecenoic acid as natural quorum sensing inhibitors of LuxS/AI-2 of Bacillus subtilis and LasI/LasR of Pseudomonas aeruginosa. Food Res Int. 2023 Mar:165:112519. [CrossRef]
- Abdulrahman, Hayder; Misba, Lama; Ahmad, Shabbir; Khan, Asad U.: Curcumin induced photodynamic therapy mediated suppression of quorum sensing pathway of Pseudomonas aeruginosa: An approach to inhibit biofilm in vitro. Photodiagnosis Photodyn Ther. 2020 Jun:30:101645. [CrossRef]
- Oh, D. H.; Marshall, D. L. (1993): Antimicrobial activity of ethanol, glycerol monolaurate or lactic acid against Listeria monocytogenes. Int J Food Microbiol. 1993 Dec;20(4):239-46. [CrossRef]
- Kabara, J. J.; Swieczkowski, D. M.; Conley, A. J.; Truant, J. P.: Fatty acids and derivatives as antimicrobial agents. Antimicrob Agents Chemother. 1972 Jul;2(1):23-8. [CrossRef]
- Sommer R, Exner TE, Titz A: A Biophysical Study with Carbohydrate Derivatives Explains the Molecular Basis of Monosaccharide Selectivity of the Pseudomonas aeruginosa Lectin LecB. PLoS One. 2014 Nov 21;9(11):e112822. [CrossRef]
- Sommer, Roman; Wagner, Stefanie; Rox, Katharina; Varrot, Annabelle; Hauck, Dirk; Wamhoff, Eike-Christian et al.: Glycomimetic, Orally Bioavailable LecB Inhibitors Block Biofilm Formation of Pseudomonas aeruginosa. J. Am. Chem. Soc. 2018, 140 (7): 2537–2545. [CrossRef]
- Kim, Bomin; Park, Ji-Su; Choi, Ha-Young; Yoon, Sang Sun; Kim, Won-Gon: Terrein is an inhibitor of quorum sensing and c-di-GMP in Pseudomonas aeruginosa: a connection between quorum sensing and c-di-GMP. Sci Rep 8, 8617 (2018). [CrossRef]
- W Auerbach, Joseph; Weinreb, Steven M.: Synthesis of terrein, a metabolite of Aspergillus terreus. J. Chem. Soc., Chem. Commun., 1974, 298-299. [CrossRef]
- Klunder, A.J.H.; Bos, W.; Zwanenburg, B.: An efficient stereospecific total synthesis of (±)-terrein. Tetrahedron Letters 1981 22 (45), 4557–4560. [CrossRef]
- Huang, Dan; Yang, Jianni; Li, Chen; Hui, Yang; Chen, Wenhao: Recent Advances in Isolation, Synthesis and Biological Evaluation of Terrein. Chem Biodivers. 2021 Dec;18(12):e2100594. [CrossRef]
- Altenbach, Hans-Josef; Holzapfel, Winfried: Synthesis of (+)-Terrein from L-Tartaric Acid. Angew Chem Int Ed Engl Vol.29, 1 Jan 1990 p. 67-68. [CrossRef]
- T Asfour, Hani Z.; Awan, Zuhier A.; Bagalagel, Alaa A.; Elfaky, Mahmoud A.; Abdelhameed, Reda F. A.; Elhady, Sameh S.: Large-Scale Production of Bioactive Terrein by Aspergillus terreus Strain S020 Isolated from the Saudi Coast of the Red Sea. Biomolecules 2019 9, 480. [CrossRef]
- Nowicki, J.; Woch, J.; Mościpan, M.; Nowakowska-Bogdan, E.: Micellar effect on the direct Fischer synthesis of alkyl glucosides. Appl Catal A: Gen 2017 539, p. 13–18. [CrossRef]
- Stickler, D. J.; Morris, N. S.; Winters, C.: Simple physical model to study formation and physiology of biofilms on urethral catheters. Methods Enzymol. 1999:310:494-50. [CrossRef]
- Loose, Maria; Naber, Kurt G.; Hu, Yanmin; Coates, Anthony; Wagenlehner, Florian: Serum bactericidal activity of colistin and azidothymidine combinations against mcr-1-positive colistin-resistant Escherichia coli. Int J Antimicrob Agents. 2018 Dec;52(6):783-789. [CrossRef]
- https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/QC/v_12.0_EUCAST_QC_tables_routine_and_extended_QC.pdf48.
- Jones, Steven M.; Yerly, Jerome; Hu, Yaoping; Ceri, Howard; Martinuzzi, Robert: Structure of Proteus mirabilis biofilms grown in artificial urine and standard laboratory media. FEMS Microbiol Lett. 2007 Mar;268(1):16-21. [CrossRef]
- Loose, Maria; Naber, Kurt G.; Purcell, Larry; Wirth, Manfred P.; Wagenlehner, Florian M. E.: Anti-Biofilm Effect of Octenidine and Polyhexanide on Uropathogenic Biofilm-Producing Bacteria. Urol Int. 2021; 105 (3-4): 278-284. [CrossRef]
- Hola, Veronika; Peroutkova, Tereza; Ruzicka, Filip: Virulence factors in Proteus bacteria from biofilm communities of catheter-associated urinary tract infections. FEMS Immunol Med Microbiol. 2012 Jul;65(2):343-9. [CrossRef]
- Sommer, Roman; Rox, Katharina; Wagner, Stefanie; Hauck, Dirk; Henrikus, Sarah S.; Newsad, Shelby et al.: Anti-biofilm Agents against Pseudomonas aeruginosa: A Structure-Activity Relationship Study of C-Glycosidic LecB Inhibitors. J Med Chem. 2019 Oct 24;62(20):9201-9216. [CrossRef]
- https://www.sigmaaldrich.com/DE/de/product/sigma/t5705?srsltid=AfmBOory9kFAwnPgePBFzgs71ECzIj_LE8GZGAo4lxziIEK9f-eytRf_.
- Kasorn A, Loison F, Kangsamaksin T, Jongrungruangchok S, Ponglikitmongkol M. Terrein inhibits migration of human breast cancer cells via inhibition of the Rho and Rac signaling pathways. Oncol Rep. 2018 Mar;39(3):1378-1386. [CrossRef]
Figure 1.
Experimental Setup. 1a) -80C glycerol stock of P. aeruginosa ATCC 27853, 1b) spread on LB-Agar with following 24h incubation, 1c) primary culture of P. aeruginosa ATCC 27853, 1d) secondary culture with three catheter-pieces in medium, 1e) dilution row made from biofilm suspension obtained from processed catheter-pieces, 1f) spread appropriate dilutions on LB agar with following 24h incubation for CFU determination.
Figure 1.
Experimental Setup. 1a) -80C glycerol stock of P. aeruginosa ATCC 27853, 1b) spread on LB-Agar with following 24h incubation, 1c) primary culture of P. aeruginosa ATCC 27853, 1d) secondary culture with three catheter-pieces in medium, 1e) dilution row made from biofilm suspension obtained from processed catheter-pieces, 1f) spread appropriate dilutions on LB agar with following 24h incubation for CFU determination.
Figure 2.
Influence of culture media and low molecular substances on the biofilm growth of P. aeruginosa ATCC 27853. Biofilms were determined via CFU-count, 1cm latex-catheter-pieces served as substrate for biofilm growth. Culture medium for tests was AU, incubation time 24 h at 37°C. 2a) different media chosen, left: artificial urine (AU), right: Lysogeny Broth-Medium (LBM), 2b) 512 µg/mL curcumin (CURC) and 5120 µg/mL Soluplus® (SOL), 2c) 512 µg/mL resveratrol (RES) and 5120 µg/mL Soluplus® (SOL ), 2d) 256 µg/mL monolaurin (ML) and 48 µg/ml polyhexanide (Prontosan®; PRT), 2e) 512 µg/ml monolaurin (ML) and 5120 µg/mL Soluplus® (SOL), degree of significance is shown using ** for p ≤ 0.01 and ***p ≤ 0.001, 2a: for statistical analysis a 2 tailed t-Test was used instead of an ANOVA – analysis.
Figure 2.
Influence of culture media and low molecular substances on the biofilm growth of P. aeruginosa ATCC 27853. Biofilms were determined via CFU-count, 1cm latex-catheter-pieces served as substrate for biofilm growth. Culture medium for tests was AU, incubation time 24 h at 37°C. 2a) different media chosen, left: artificial urine (AU), right: Lysogeny Broth-Medium (LBM), 2b) 512 µg/mL curcumin (CURC) and 5120 µg/mL Soluplus® (SOL), 2c) 512 µg/mL resveratrol (RES) and 5120 µg/mL Soluplus® (SOL ), 2d) 256 µg/mL monolaurin (ML) and 48 µg/ml polyhexanide (Prontosan®; PRT), 2e) 512 µg/ml monolaurin (ML) and 5120 µg/mL Soluplus® (SOL), degree of significance is shown using ** for p ≤ 0.01 and ***p ≤ 0.001, 2a: for statistical analysis a 2 tailed t-Test was used instead of an ANOVA – analysis.
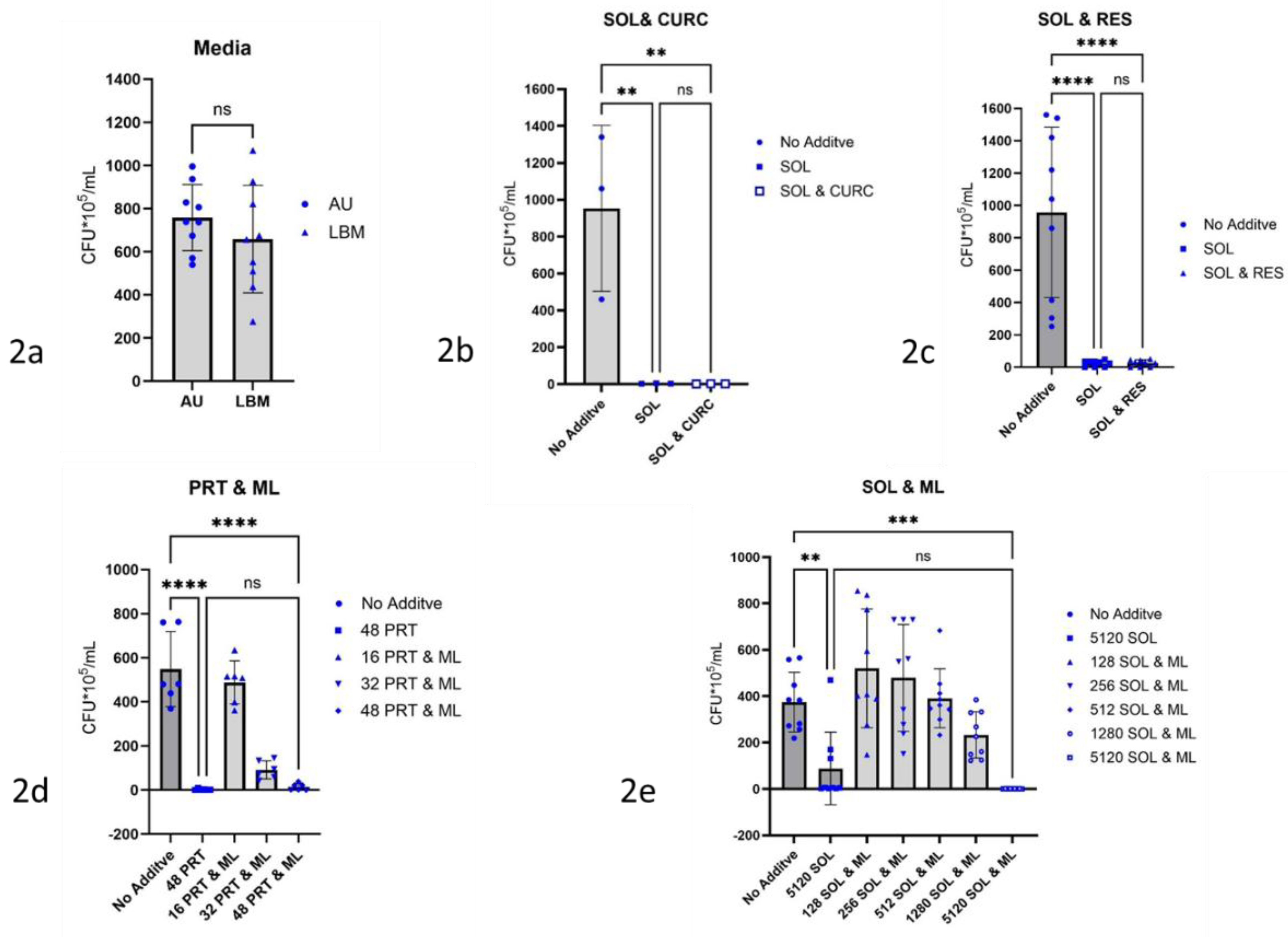
Figure 3.
Glycomimetics as inhibitors of biofilm growth of P. aeruginosa ATCC 27853. Biofilms were determined via CFU-count, 1 cm latex-catheter-pieces served as substrate for biofilm growth. Culture medium for tests was AU, incubation time 24h at 37 °C. 3a) n-undecyl-α-d-mannopyranoside (nU-Man) in different amounts µg/mL, 3b) n-undecyl-α/β-l-fucopyranoside (nU-Fuc) in different amounts µg/mL, significance is shown using * for p ≤ 0.05 and *** for p ≤ 0.001, 3c) n-undecyl-α/β-l-fucopyranoside (nU-Fuc) in 32 – 512 µg/mL as inhibitor of biofilm growth of P. aeruginosa PA01.
Figure 3.
Glycomimetics as inhibitors of biofilm growth of P. aeruginosa ATCC 27853. Biofilms were determined via CFU-count, 1 cm latex-catheter-pieces served as substrate for biofilm growth. Culture medium for tests was AU, incubation time 24h at 37 °C. 3a) n-undecyl-α-d-mannopyranoside (nU-Man) in different amounts µg/mL, 3b) n-undecyl-α/β-l-fucopyranoside (nU-Fuc) in different amounts µg/mL, significance is shown using * for p ≤ 0.05 and *** for p ≤ 0.001, 3c) n-undecyl-α/β-l-fucopyranoside (nU-Fuc) in 32 – 512 µg/mL as inhibitor of biofilm growth of P. aeruginosa PA01.
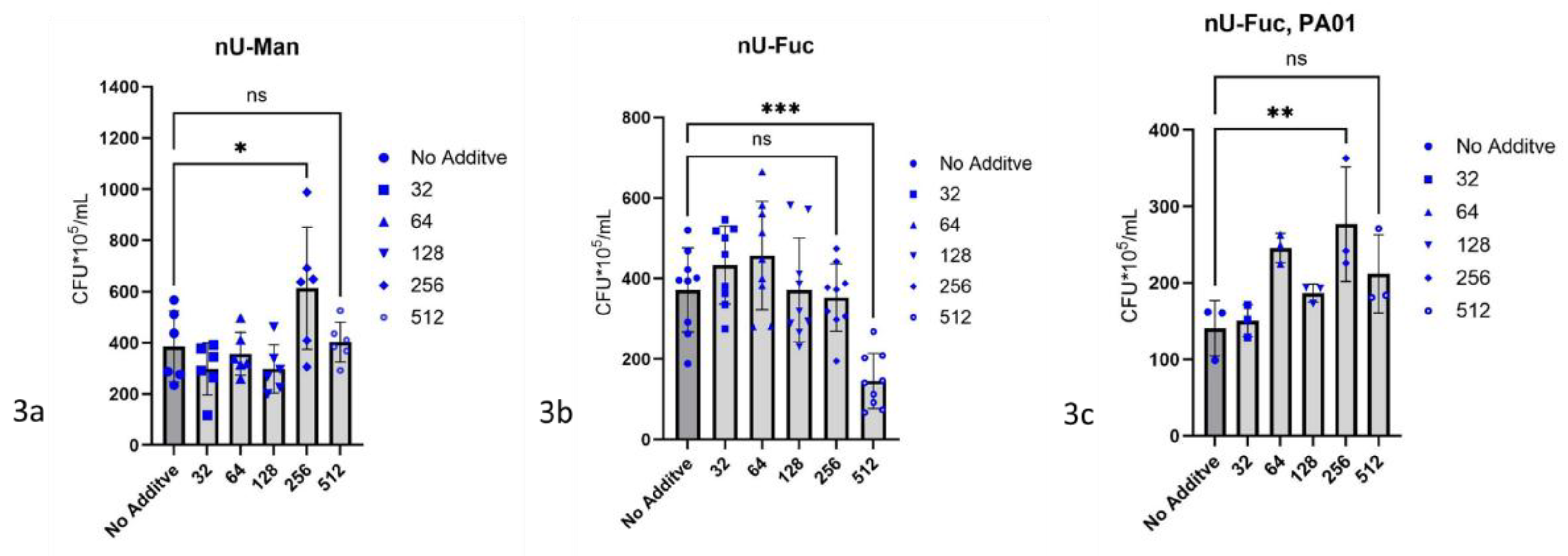
Figure 4.
Influence of terrein on the biofilm growth of P. aeruginosa ATCC 27853. Biofilms were determined via CFU-count, 1cm latex-catheter-pieces served as substrate for biofilm growth. Culture medium for tests was AU, incubation time 24 h at 37°C. 4a) Terrein (TER) in different concentrations µg/mL, 4b) 256 µg/ml terrein and 64 µg/mL polyhexanide (Prontosan®; PRT), 4c) molecular structure of terrein with terrein producing Aspergillus terreus SO22 (photo from Asfour et al., 2019), degree of significance is shown using * for p ≤ 0.05, *** p ≤ 0.001, and ****p ≤ 0.0001.
Figure 4.
Influence of terrein on the biofilm growth of P. aeruginosa ATCC 27853. Biofilms were determined via CFU-count, 1cm latex-catheter-pieces served as substrate for biofilm growth. Culture medium for tests was AU, incubation time 24 h at 37°C. 4a) Terrein (TER) in different concentrations µg/mL, 4b) 256 µg/ml terrein and 64 µg/mL polyhexanide (Prontosan®; PRT), 4c) molecular structure of terrein with terrein producing Aspergillus terreus SO22 (photo from Asfour et al., 2019), degree of significance is shown using * for p ≤ 0.05, *** p ≤ 0.001, and ****p ≤ 0.0001.
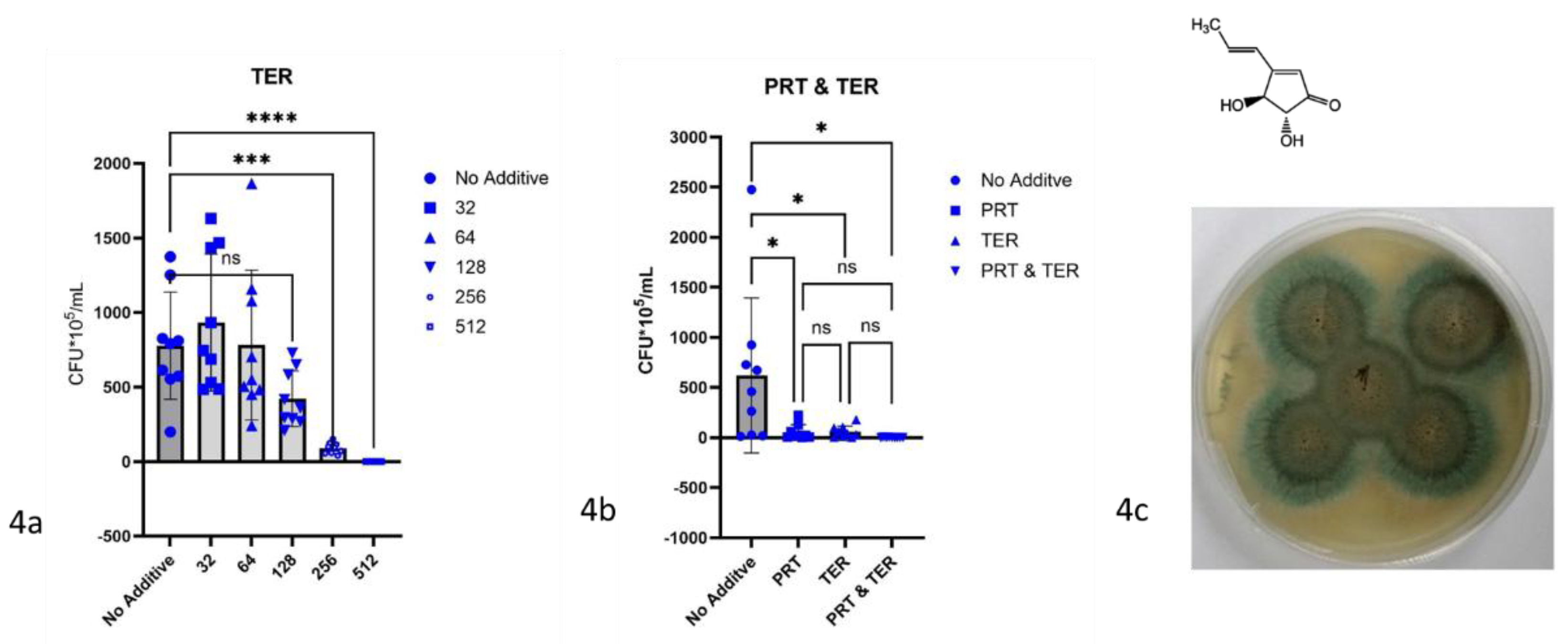

Table 1.
Overview of substances tested for inhibition of biofilm formation by P. aeruginosa.
| Substance | Abbreviation | Literature | Vendor |
| Curcumin & Soluplus | CURC & SOL | ||
| Curcumin | CURC | Abdulrahman et al. 2020 [33] | Merck Millipore # CAS 458-37-7 |
| Resveratrol & Soluplus | RES & SOL | ||
| Resveratrol | RES | Qi et al. 2022 [28] | TCI # CAS RN®: 501-36-0 |
| Soluplus | SOL | Shamma and Basha 2013 [18] |
BASF SE Ludwigshafen am Rhein Germany |
| Monolaurin | ML | Oh and Marshall 1993 [34]; Kabara et al. 1972 [35] | TCI # CAS RN®: 142-18-7 |
| Monolaurin & Soluplus | ML & SOL | ||
| Prontosan | PRT | Loose et al. 2021[50] | B. Braun Melsungen AG, Melsungen, Germany |
| Terrein | TER | Kim et al. 2018 [38] | AdipoGen # CAS-No. 582-46-7 |
| Terrein & Prontosan | TER & PRT | ||
| n-Undecyl-α-D-Mannopyranoside | nU-Man | Nowicki et al. 2017 [44]; Sommer et al. 2018 [37] |
laboratory synthesis |
| n-Undecyl-α/β-L-Fucopyranoside | nU-Fuc | Nowicki et al. 2017 [44]; Sommer et al. 2018 [37] |
laboratory synthesis |
| Piperacillin & Soluplus | PIP & SOL | ||
| Piperacillin | PIP | EUCAST QC Tables [48] | Sigma-Aldrich # CAS-No. 66258-76-2 |
Table 2.
MIC-Values P. aeruginosa.
| Substance | MIC in AU (P. aeruginosa ATCC 27853) |
| Curcumin & Soluplus | > 512 & > 5,120 |
| Curcumin | not determined |
| Resveratrol & Soluplus | > 512 & > 5,120 |
| Resveratrol | > 512 |
| Soluplus | > 5120 |
| Monolaurin | > 512 |
| Monolaurin & Soluplus | |
| Prontosan | 32/ 32 |
| Terrein | 512 |
| Terrein & Prontosan | 128 & 32/16 |
| n-Undecyl-α-D-Mannopyranoside | not determined |
| n-Undecyl-α/β-L-Fucopyranoside | not determined |
| Piperacillin & Soluplus | 4 & 40 |
| Piperacillin | 4 |
Note: Minimal inhibitory concentration (MIC)-Values of n-undecyl-α-d-mannopyranoside and n-undecyl-α/β-l-fucopyranoside are not determined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



