
Submitted:
24 October 2024
Posted:
25 October 2024
You are already at the latest version
Abstract
Plants of the subfamily Amaryllidoideae are a source of unique and bioactive alkaloids called Amaryllidaceae alkaloids. The study of their anticancer potential has intensified in recent years. This work aims to locate and characterize the profile of cytotoxic alkaloids biosynthesized and stored in different tissues of Phaedranassa lehmannii Regel using different histochemical methods and chromatographic analysis. The histochemical analysis in the bulbs revealed the presence of alkaloids at the basal edge of scale-like leaves and bud apical zone. The GC-MS analysis indicated that the bulbs biosynthesize alkaloids distributed in crinine- (9.80 µg/g DW), galanthamine- (8.04 µg/g DW), lycorine- (7.38 µg/g DW) and narciclasine-type (3.75 µg/g DW). The root biosynthesizes alkaloids distributed mostly in lycorine- (225.29 µg/g DW) and galanthamine-type (0.723 µg/g DW) alkaloids. The total alkaloids biosynthesized by the root (324.93 µg/g DW) exceeded eleven times the abundance of the alkaloids identified in the bulbs (28.97 µg/g DW). In addition, the total alkaloids fractions exhibited a cytotoxic effect dose‐dependent in the evaluated concentrations with IC50 values of 11.76 ± 0.99 µg/mL and 2.59 ± 0.56 µg/mL against human lung (A549) cancer cells and 8.00 ± 1.35 µg/mL and 18.74 ± 1.99 µg/mL against gastric (AGS) cancer cells. The present study provided evidence to locate and characterize the alkaloids of P. lehmannii grown under nursery conditions as a species producing potential antiproliferative alkaloids.
Keywords:
Phaedranassa lehmannii
; Amaryllidaceae alkaloids
; cytotoxicity
; histochemical localization.
1. Introduction
The family Amaryllidaceae is a monophyletic group, represented by approximately 1650 species distributed in 73 genera. Three clades are recognized, represented in the subfamilies Agapanthoideae, Allioideae, and Amaryllidoideae. The subfamily Amaryllidoideae is made up of monocotyledonous, bulbous, and flowering species [1]. The subfamily Amaryllidoideae has increased its level of importance due to the presence of unique isoquinoline alkaloids biogenetically derived from tyrosine and phenylalanine, and known as Amaryllidaceae alkaloids [2,3]. These alkaloids are characterized by their biological effects such as neuroprotective, enzyme inhibitors of cholinesterase and butyrylcholinesterase, anti-inflammatory, and antiviral, among others [2]. Galanthamine has demonstrated a superior response in clinical terms in comparison with other Amaryllidaceae alkaloids [4], and is approved for the treatment of Alzheimer's disease [5]. On the other hand, Amaryllidaceae alkaloids have been isolated and tested also as anti-proliferative agents for different types of cancer [6,7]. The biological potential shows that Amaryllidaceae alkaloids could be used in the pharmacological field of cancer [1,8].
In Colombia, 18 genera and 48 species of the Amaryllidoideae subfamily have been reported, of which 11 genera and 27 species are native with 9 endemic species [9]. Phaedranassa, one of these genera, correspond to perennial herbs, up to 60 cm tall, with bulbs 2-6 cm in diameter, globose, and with cream to brown tunic. The species of the genus are distributed in tropical and premontane forests of the southern Cordillera Occidental and Nudo de los Pastos, reaching the center of Ecuador [9]. Although phytochemical analysis of Phaedranassa have shown three different Amaryllidaceae alkaloid types (lycorine, crinine/haemanthamine, and galanthamine), the alkaloids identified vary according to the species [10,11]. Phaedranassa cinerea, Phaedranassa cuencana, and Phaedranassa ventricosa are characterized by producing lycorine- and galanthamine-type alkaloids [7,10]. In Phaedranassa brevifolia, Phaedranassa dubia, Phaedranassa glauciflora and Phaedranassa tunguraguae, the presence of crinine/haemanthamine-, lycorine- and galanthamine-type alkaloids is notable [10,11,12]. In addition, alkaloids such as crinine-, lycorine- and narcyclasine-type, have been reported in Phaedranassa lehmannii [13,14]. Other less frequent alkaloid types are homolycorine and montanine, which were found in fewer species [10,12]. In this way, the production of crinine- and lycorine-type alkaloids in Phaedranassa species is highlighted.
In the search for new molecules for the treatment and prevention of cancer, the vegetative organs of many Amaryllidoideae species have been analyzed as a source of numerous cytotoxic Amaryllidaceae alkaloids, particularly crinine- and lycorine-type alkaloids [15,16]. It has been suggested the correlation between the cytotoxic effect of Amaryllidaceae species and crinine- and lycorine-type alkaloids [7]. Simultaneously, the location, the composition of the chemical profile, and the distribution of alkaloids in producing organs are considered important productive features of the subfamily Amaryllidoideae, due to the quantity of compounds generally in low concentrations [2]. Histochemistry identification may yield valuable data on the producing organs and cell-type localization of the Amaryllidaceae alkaloids [17]. For this, different histochemical methods have been developed, which allow the characterization of plant structures where different compounds are distributed and accumulate, as well as the precise determination of tissues involved in the secretion of alkaloids [18]. However, studies of spatial distribution, localization, and accumulation of Amaryllidaceae alkaloids in different plant tissues (bulb, leaf, and root) are limited, especially in cytotoxic Amaryllidaceae alkaloids. In most cases, and for industrial purposes, histochemical studies have been performed on the main sources of galanthamine, and alternative sources of galanthamine for further optimization of compound production [17,19]. Therefore, this work aims to locate and characterize the alkaloids synthesized and stored by different histochemical methods and chromatographic analysis of plants grown under nursery conditions of P. lehmannii, a species with interesting pharmacological properties and a source of cytotoxic alkaloids [7].
2. Results
2.1. Histochemical Analysis
P. lehmannii bulbs develop a thin tunic and three scale-like leaves that protect the bud (Figure 1A). Scales leaves are made up of thin-walled polyhedric parenchyma cells. The scaly leaf closest to the bud develops lysogenic cavities in the apical zone (Figure 1E). In addition, the bulbs present a basal plate formed by vascular tissue (xylem vessels) and parenchyma flattened cells, which have amyloplasts storage (Figure 1B,D), and a central bud provided by a base formed by parenchyma-like storage cells, while the bud apical zone has developing vascular tissue (Figure 1C). Dragendorff's and Wagner's reagents with reddish-brown staining, and Lugol's reagent brown, showed alkaloid storage at the basal edge of scale-like leaves and the bud apical zone with the absence of alkaloids on the basal plate (Figure 1F–J).
2.2. Chromatographic Analysis of Alkaloids from P. lehmannii
After histochemical and phytochemical detection with different staining reagents, the alkaloids present in the bulb and root of P. lehmannii were analyzed by GC-MS. In the total alkaloid fraction of bulbs, eight alkaloids were detected with a predominant abundance of crinine- (9.80 µg/g DW), followed by galanthamine- (8.04 µg/g DW) and lycorine-type (7.38 µg/g DW) alkaloids (Figure 2, Table S1). The alkaloids 8-O-demethylmaritidine (8.77 µg/g DW) and lycorine (6.79 µg /g DW) were the most abundant. Interestingly, the root has biosynthesis of nine alkaloids with high diversity and abundance. Lycorine-type alkaloids (225.29 µg/g DW) predominate in the root, with a high prevalence of anhydrolycorine (133.7 µg/ g DW) and lycorine (73.79 µg/ g DW), being the most abundant compound in the total alkaloid fraction, followed by galanthamine-type (72.35 µg/g DW) and narciclasine- type (13.74 µg/g DW) alkaloids. Additionally, the results showed that the alkaloids biosynthesized by the root (324.93 µg /g DW) exceed eleven times the abundance of the alkaloids identified in the bulbs (28.97 µg/g DW).
2.3. Cytotoxic Activity from P. lehmannii alkaloids
The alkaloids fraction of bulbs and roots showed a cytotoxic effect in the evaluated concentrations with IC50 values of 11.76 ± 0.99 µg/mL and 2.59 ± 0.56 µg/mL against human lung (A549) cancer cells, and 8.00 ± 1.35 µg/mL and 18.74 ± 1.99 µg/mL against gastric (AGS) cancer cells for 48 h exposure, determined by the reduction of 3-(4,5-dimethylthiazol-2) 2,5- diphenyltetrazolium bromide (MTT) to formazan. The alkaloids showed a concentration-dependently decreased in cell viability with statistically significant differences for concentrations greater than 1 μg/mL (Figure 3A,B). Lycorine and doxorubicin were tested as standards (positive control) with IC50 values of 4.97 ± 0.89 µg/mL and 5.52 ± 0.21 µg/mL, respectively (Table 1). Therefore, the values showed that alkaloids biosynthesized in the root generate a high cytotoxic effect on lung cancer cells, compared to lycorine. Alkaloid fractions were also evaluated on human keratinocytes (HaCat, non-cancer control cells) to evaluate the degree of selectivity of the alkaloids by determination of the selectivity index (SI, ratio of IC50). The SI values higher than 1.0 indicate that compounds or extracts have anticancer specificity. If SI values are much greater than 1.0, the samples are highly selective [20]. Based on this, the SI data shown in Table 1 indicate that alkaloids of bulbs, root, and lycorine standard exhibit a high degree of cytotoxic selectivity against lung and gastric cancer cells with values between 2.11 and 15.29.
3. Discussion
Alkaloids are molecules that plants use for chemical defense. Most plants produce alkaloids in one organ, then transport them through the xylem or phloem and store in organs such as seeds, flowers, leaves, or more specialized storage organs such as bulbs [21]. In addition, it is well known that the production of metabolites depends on conditions such as the state of development of the plant, and environmental conditions such as climate, location, and method of cultivation [22]. Specifically in the genus Phaedranassa, a chemical diversity in the production of Amaryllidaceae alkaloid has been demonstrated [10-14], and it could be suggested that the production is regulated by the processes of alkaloid biosynthesis in specific tissues or cells, depending largely on the level of differentiation and development, and then stored mainly in bulbs and roots, as occurs in other species of the subfamily [17]. However, few studies have examined the anatomy and histochemistry of Phaedranassa bulbs. In most cases, histochemical studies have been performed on the main sources of galanthamine for further optimization of the compound production [17,19].
In Amaryllidoideae species, a previous study to identify the alkaloids produced by Rhodophiala bifida highlights alkaloid concentrations in bulbs and roots [3]. While in Narcissus pseudonarcissus it has reported high alkaloid (galanthamine) storage in basal plate followed by bulb tissues [23]. Research on Zephyranthes irwiniana bulbs identified alkaloids in the chlorenchyma cells of the leaf and bulb margins and reported that data on the histolocalization of Amaryllidaceae alkaloids are scarce [24]. However, our results show that alkaloid storage in P. lehmannii occurs both in the roots and in the bulbs, in the latter, in the scale-like leaves, and in the apical zone of the buds, but not in the basal plate (Figure 1). This confirms the possible movement of alkaloids between tissues suggested by Amaryllidoideae species [25]. Nevertheless, further studies are required to characterize the transport of cytotoxic alkaloids and identify in which tissues their synthesis and storage occur. In this regard, it has been shown in Hippeastrum papilio that galantamine biosynthesis occurs in green tissues and is transferred to other plant organs, for example, bulbs and roots [17].
The results of the localization of alkaloids in plant tissues could provide a better understanding of the role and sites of biosynthesis and accumulation of these secondary metabolites. In addition, this contributes to the search for new sources and to the establishment of organogenesis processes aimed at the production of alkaloid-rich biomass, without endangering the biodiversity of wild populations [26,27]. It would be a strategy to increase the availability of cytotoxic alkaloids with interesting pharmacological activities such as lycorine, limited by nature in plants of the Amaryllidoideae subfamily [28,29].
In the analysis of the identification of alkaloids in bulbs and roots from P. lehmannii, it was established that the alkaloids detected by CG-MS would be directly correlated with the positive staining observed for alkaloids accumulated in the central part of the root, and at the basal edge of scale-like leaves and in the bud apical zone, these two lasts of the bulbs. The fraction of total alkaloids of bulbs corresponded to a relative abundance of 28.97 µg /g DW with a high prevalence of crinane-, galanthamine- and lycorine-type alkaloids, with an outstanding abundance of 8-O-demethylmaritidine, galanthamine, saguinine, and lycorine (Figure 2). Complementary, the production of alkaloids in the root (324.93 µg /g DW) was eleven times higher than that reported for bulbs. In addition, in the root predominant the lycorine-type alkaloids, mainly anhydrolycorine and lycorine, followed by galantamine-type alkaloids, with a predominance of galantamine. This shows that biosynthesis and accumulation of specific alkaloids can be limited to specific cell types, tissues, or organs [30,31].
This effect has been observed in Amaryllidoideae species such as Lycoris radiata, where leaves and roots contain significantly more alkaloids of interest than in scapes and bulbs [31]. In Leucojum aestivum, the biosynthesis and storage of galantamine is higher in bulb (0.0949 mg/g) than in root (0.0262 mg/g), while lycorine is higher in root (0.2328 mg/g) than bulb (0.1994 mg/g) [29]. Similarly, Galanthus nivalis produces twenty-two times more lycorine in root (0.1816 mg/g) than in bulb (0.0080 mg/g) [29,32]. Therefore, the results found for P. lehmannii are related to those reported for other plants of the subfamily Amarylllidoideae regarding the abundance of alkaloids such as lycorine and galanthamine. Interestingly, these two alkaloids are widely required by the pharmaceutical industry due to insufficient availability of natural resources and the significant increase in the demand for galanthamine for the treatment of neurodegenerative diseases [28] and biological interest of lycorine for the search of therapeutic alternatives in cancer [33,34].
Although the alkaloids identified in P. lehmannii have been reported in previous studies [7,8,13,14], differences in the concentration of alkaloids and the presence of others previously not reported, such as sternbergine, were evidenced. This is possibly due to a change in some biological, chemical, or environmental factor that influences the biosynthesis and accumulation of secondary metabolites [35]. Concerning other species of the genus, 70% of the alkaloids identified in P. dubia and P. brevifolia correspond to lycorine- and haemanthamine/crinine-type alkaloids [12]. In P. cinerea, P. cuencana, P. dubia, P. glauciflora and P. tunguraguae approximately 50% of the alkaloids correspond to the lycorine-type and 15% to the crinine/hemanthamine-type alkaloids [10]. Therefore, plants of the genus Phaedrannassa are chemically interesting for their alkaloid profiles, and pharmacologically of interest due to the cytotoxic potential that many of these alkaloids present [12].
The alkaloid fractions from P. lehmannii were evaluated in gastric (AGS) and lung (A549) cancer cells using lycorine as the reference alkaloid. The alkaloid fraction of the root showed a high cytotoxic effect on lung cancer cells with IC50 values of 2.59 ± 0.56 µg/mL, compared to the alkaloid fraction of the bulbs. The antiproliferative effect against lung cancer cells could be attributed to the high presence of lycorine-type alkaloids in the root (225.29 µg/g DW), every time that lycorine induces high cell death with IC50 of 4.07 ± 0.21 µg/mL, with a high degree of selectivity. It has been previously reported that lycorine-type alkaloids and lycorine have cytotoxic efficacy against many cancers cell lines, including lung cancer [34,36,37,38]. Lycorine is a potent inducer of apoptosis, both in mitochondrial and death receptor-mediated apoptosis. This mode of action is related to the inhibition of migration and proliferation of cells with resistance to apoptosis [36]. Although multiple apoptotic mechanisms of lycorine are possible, depending on the type of cancer, in human lung cancer cells lycorine causes a G0/G1 phase arrest, induces an early stage of apoptosis and initiates mitochondrial dysfunction [38]. Therefore, lycorine has been suggested as an excellent candidate for combating cancers [34,36], and new lycorine derivatives have been synthesized to verify the antiproliferative effects against different cancer cell lines [39].
4. Materials and Methods
4.1. Plant Material
The plant material of the plant grown under greenhouse conditions was obtained from the Plant Biotechnology Unit, Faculty of Engineering, Universidad Católica de Oriente. Rionegro, Colombia. One specimen of the species was deposited in the Herbarium of the University of Antioquia, Medellín, Colombia (voucher 5106). The material studied was collected with authorization from the Ministry of Environment with a genetic resource access contract #328. The selection criteria were material not in bloom and not more than 20 cm in length.
4.2. Histochemical Analysis
Bulbs of P. lehmannii were fixated in formaldehyde, acetic acid, and ethanol (FAA) for 24 hours at 6°C [40]. After, each tissue was dehydrated in an ethanol series (50, 60, 70. 80, 90, 96 and 100%) and rinsed in HistoChoice® twice for 12 hours. Subsequently, the tissues were embedded in a solution 2:1 composed of HistoChoice® and Paraplast® paraffin for 2 hours and then in a 1:2 solution. Finally, the tissues were embedded in 100% Paraplast® paraffin for 12 hours for three times [41]. Sections were made in different cutting planes of 5 a 7 µm thick, using a Leica RM2125 rotatory microtome. Staining was done with Safranin-Alcian Blue (Fasga) for structural differentiation of tissues [42]. Dragendorff and Wagner methods were used for the detection of alkaloids [43]. With Dragendorff reagents, the alkaloids were stained red or red-brown. In Wagner assay, the reagent gives colors similar to Dragendorff in the presence of alkaloids. Lugol's reagent, which is similar in chemical composition to Wagner reagent, produces a golden-brown reaction [43,44,45]. Sections were observed through an Olympus CX31 optical microscope.
4.3. Extraction of Alkaloids
The treatment of plant material and extraction of alkaloids was carried out following protocols described previously [7,46]. The plants grown under greenhouse conditions were collected and washed with drinking water. After that, the bulbs were cut into pieces of 3 cm and dried to 40°C for 48 hours. The extract in methanol was made with dried and ground plant material, applying 15 min of ultrasonic baths (3 times), changing the solvent and evaporating the solvent at reduced pressure. The concentrated extract was dissolved in 10 mL 2% H2SO4, hexane (3x10 mL) was used to remove neutral compounds and ammonium hydroxide was added to the aqueous fraction to adjust pH in the range of 9 to 10. Then, the alkaloids were extracted with chloroform (3 x 50mL). The organic solvent was evaporated using a rotary evaporator. To finish, for analysis in GC-MS 5mg of extract was dissolved in 500 μL methanol.
4.4. Chromatographic Analysis of Alkaloids
The analysis of alkaloids of P. lehmannii by GC-MS was performed according to the method described previously [47]. The alkaloid fraction was injected into an Agilent 7890 Gas Chromatograph equipped with a 5975C selective mass detector and electronic impact (EI) operating to 70 eV in splitless mode, programmed to acquire signals in scan mode between 40 and 400 daltons. In an HP-1 MS capillary column (30m x 0.25 mm x 0.25μm) were separated the alkaloids. It has a carrier gas flow (Helium) of 1 mL/min. The temperature ramp was as follows: 100-180°C at 15°C/minute, 180-300°C at 5°C/minute, and 10 minutes hot at 300°C. The temperature of the injector is kept at 250°C. Subsequently, 1 μL of the sample was injected.
4.5. Determination of the Alkaloid Profile
To identify alkaloids, the database “Amaryllidaceae Alkaloid Spectroteca, Agro Bio Institute (Bulgaria)” and data reported in scientific journals were used, to compare the fragments of the mass spectra of each molecule. The Kovats retention rates of the compounds were recorded with a standard calibration mixture of n-hydrocarbons (C7-C40). The percentage TIC (total ion current) was determined for each alkaloid. The codeine (50 μg/mL) was used as an internal standard and with this, the abundance of each compound was calculated. The proportion of each compound in the extracts tested was expressed as a percentage of the total alkaloid content the area of GC-MS peaks depends on the concentration of the related compounds and the intensity of their mass spectral fragmentation.
4.6. Cell Viability of Alkaloid Fraction
Cell culture: Cell viability of the total alkaloids fraction was evaluated in the gastric (AGS, CRL-1739™) and lung (A549, CCL-185™) cancer cell line. Human keratinocytes (HaCat, PCS-200-011™) were used as a non-carcinogenic control cell line. The cells A549 and HaCat were grown cultured in Dulbecco Modified Eagle Medium-high glucose (DMEM) and AGS in Ham's F-12 nutrient medium (F12) supplemented with 10% heat-inactivated FBS, 100 U/mL penicillin and 100 μg/mL streptomycin, 2 mM L-glutamine in a humidified atmosphere of 5% CO2 and 95% air at 37°C. Cells were monitored on a Nikon Eclipse TS100 inverted phase contrast microscope. Cell viability experiments were performed when cells reached 75-80% confluence using 0.25% Trypsin 1 mM EDTA.
Cell viability by mitochondrial MTT reduction: Cell viability was determined using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The total alkaloids fraction and lycorine standard were evaluated at concentrations of 50, 30, 20, 10, 1, 0.1 μg/mL and doxorubicin at 10, 5, 2.50, 1.25, 0.62, 0.31 μg/mL. All samples were prepared in dimethyl sulfoxide (DMSO). Cells were seeded in 96-well plates with a cell density of 2 x 106 cells/mL, incubated for 24 h at 37 °C with 5% CO2. After this time, cells were incubated with samples at 37 °C with 5% CO2 for 48 h. Subsequently, 50 μL of MTT (1.0 mg/mL buffered saline phosphate) was added. After, 4 h of incubation at 37 °C, the MTT culture medium was removed and replaced with 150 μL DMSO to dissolve the formazan crystals. Plates were incubated in the dark with agitation for 2 h. Optical density was determined at 540 nm using a microplate reader. All experiments were conducted with three independent trials, each with six replicas.
4.7. Statistical Analysis
The results are shown as the mean ± SD. The statistical significance between the control group and the treatments with the alkaloid fraction was evaluated by a one-way analysis of variance (ANOVA) followed by the Dunnett multiple-comparison test, using the GraphPad Prism 5.0 data analysis system.
5. Conclusions
In summary, this study provides relevant information about the histochemical localization of alkaloids of Amaryllidaceae in the roots, and in the basal edge of scale-like leaves and bud apical zone of P. lehmannii. Histochemical analysis of plants grown under nursery conditions of P. lehmannii showed that a possible site of alkaloid biosynthesis is the basal edge of scale-like leaves and the apical zone of buds. Where, lycorine-type alkaloids such as anhydrolycorine prevalence at root, and crinine-type alkaloids such as 8-O-demethylmaritidine in bulbs. Therefore, the roots of P. lehmannii represent a good source of lycorine. The bulbs constitute a significant part of the plant biomass and contain high levels of crinine-type alkaloids, not detected in the root, so they could also be considered as a valuable source of alkaloids. The cytotoxic potential against lung cancer cells of the root alkaloids was reported, mainly attributed to the presence of lycorine-type alkaloids. Consequently, the underground organs (bulb and root) of P. lehmannii are a source of alkaloids with cytotoxic potential, with interesting pharmacological properties in lung and gastric cancer.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Amaryllidaceae alkaloids identified in Phaedranassa lehmannii by GC–MS.
Author Contributions
Conceptualization, E.O. and D.C.; methodology, L.M.T., H.L., J.M.R., I.Z.V. and M.D; formal analysis, L.M.T. and I.Z.V.; investigation, L.M.T., H.L., J.M.R., I.Z.V. and M.D.; resources, E.O. and D.C.; writing—original draft preparation, E.O., D.C., L.T. and I.Z.V.; writing—review and editing, E.O., D.C. and L.M.T.; funding acquisition, E.O. and D.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MINCIENCIAS (Agreement # 614-2018). The authors are especially grateful to the University of Antioquia (UdeA), and the Catholic University of the East (UCO for its abbreviation in Spanish), for their important contribution to the development of this work.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the research department of Universidad Católica de Oriente for supporting the activities carried out for this work.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the study's design; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Cahlíková, L.; Kawano, I.; Řezáčová, M.; Blunden, G.; Hulcová, D.; Havelek, R. The Amaryllidaceae alkaloids haemanthamine, haemanthidine and their semisynthetic derivatives as potential drugs. Phytochem. Rev. 2021, 20, 303–323. [Google Scholar] [CrossRef]
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chemodiversity, chemotaxonomy and chemoecology of Amaryllidaceae alkaloids. In The Alkaloids Chemistry and Biology, Cordell, G.A., Ed.; Elsevier-Academic Press: New York, EEUU, 2020; Volume 83, pp. 113–185. [Google Scholar] [CrossRef]
- Reis, A.; Magne, K.; Massot, S.; Tallini, L.R.; Scopel, M.; Bastida, J.; Ratet, P.; Zuanazzi, J.A.S. Amaryllidaceae alkaloids: identification and partial characterization of montanine production in Rhodophiala bifida plant. Sci. Rep. 2019, 9, 8471. [Google Scholar] [CrossRef] [PubMed]
- Sierra, K.; de Andrade, J.P.; Tallini, L.R.; Osorio, E.H.; Yañéz, O.; Osorio, M.I.; Oleas, N.H.; García-Beltrán, O.; Borges, W.; Bastida, J.; Osorio, E.; Cortes, N. In vitro and in silico analysis of galanthine from Zephyranthes carinata as an inhibitor of acetylcholinesterase. Biomed. Pharmacother. 2022, 150, 113016. [Google Scholar] [CrossRef]
- Heinrich, M. Chapter 4 – Galanthamine from Galanthus and other amaryllidaceae – chemistry and biology based on traditional use. In The Alkaloids Chemistry and Biology; Cordell, G.A., Ed.; Elsevier-Academic Press: New York, EEUU, 2010; Volume 68, pp. 157–165. [Google Scholar] [CrossRef]
- Havelek, R.; Muthna, D.; Tomsik, P.; Kralovec, K.; Seifrtova, M.; Cahlikova, L.; Hostalkova, A.; Safratova, M.; Perwein, M.; Cermakova, E.; Rezacova, M. Anticancer potential of Amaryllidaceae alkaloids evaluated by screening with a panel of human cells, real-time cellular analysis and Ehrlich tumor-bearing mice. Chem. Biol. Interact. 2017, 275, 121–132. [Google Scholar] [CrossRef]
- Trujillo, L.; Bedoya, J.; Cortés, N.; Osorio, E.H.; Gallego, J.C.; Leiva, H.; Castro, D.; Osorio, E. Cytotoxic activity of Amaryllidaceae plants against cancer cells: biotechnological, in vitro, and in silico approaches. Molecules. 2023, 28, 260. [Google Scholar] [CrossRef]
- Trujillo-Chacón, L.M.; Alarcón-Enos, J.E.; Céspedes-Acuña, C.L.; Bustamante, L.; Baeza, M.; López, M.G.; Fernández-Mendívil, C.; Cabezas, F.; Pastene-Navarrete, E.R. Neuroprotective activity of isoquinoline alkaloids from of Chilean Amaryllidaceae plants against oxidative stress-induced cytotoxicity on human neuroblastoma SH-SY5Y cells and mouse hippocampal slice culture. Food Chem. Toxicol. 2019, 132, 110665. [Google Scholar] [CrossRef]
- Alzate, F.; Lesmes, M.; Cortés, N.; Varela, S.; Osorio, E. Sinopsis de la familia Amaryllidaceae en Colombia. Biota Colombiana. 2019, 20, 2–20. [Google Scholar] [CrossRef]
- Moreno, R.; Tallini, L.R.; Salazar, C.; Osorio, E.H.; Montero, E.; Bastida, J.; Oleas, N.H.; Acosta León, K. Chemical profiling and cholinesterase inhibitory activity of five Phaedranassa Herb. (Amaryllidaceae) species from Ecuador. Molecules 2020, 25, 2092. [Google Scholar] [CrossRef]
- Osorio, E.; Berkov, S.; Brun, R.; Codina, C.; Viladomat, F.; Cabezas, F.; Bastida, J. In vitro antiprotozoal activity of alkaloids from Phaedranassa dubia (Amaryllidaceae). Phytochem. Lett. 2010, 3, 161–163. [Google Scholar] [CrossRef]
- León, K.A.; Inca, A.; Tallini, L.R.; Osorio, E.H.; Robles, J.; Bastida, J.; Oleas, N.H. Alkaloids of Phaedranassa dubia (Kunth) J.F. Macbr. and Phaedranassa brevifolia Meerow (Amaryllidaceae) from Ecuador and its cholinesterase-inhibitory activity. S. Afr. J. Bot. 2021, 136, 91–99. [Google Scholar] [CrossRef]
- Cortes, N.; Castañeda, C.; Osorio, E.H.; Cardona-Gomez, G.P.; Osorio, E. Amaryllidaceae alkaloids as agents with protective effects against oxidative neural cell injury. Life Sci. 2018, 203, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Cortes, N.; Sierra, K.; Alzate, F.; Osorio, E.H.; Osorio, E. Alkaloids of Amaryllidaceae as inhibitors of cholinesterases (AChEs and BChEs): An integrated bioguided study. Phytochem. Anal. 2018, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.J.; Bastida, J.; Viladomat, F.; van Staden, J. Cytotoxic agents of the crinane series of Amaryllidaceae alkaloids. Nat. Prod. Commun. 2012, 7, 1677–1688. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.J.; van Staden, J. Cytotoxicity studies of lycorine alkaloids of the Amaryllidaceae. Nat. Prod. Commun. 2014, 9, 1193–1210. [Google Scholar] [CrossRef]
- Haist, G.; Sidjimova, B.; Yankova-Tsvetkova, E.; Nikolova, M.; Denev, R.; Semerdjieva, I.; Bastida, J.; Berkov, S. Metabolite profiling and histochemical localization of alkaloids in Hippeastrum papilio (Ravena) van Scheepen. J. Plant Physiol. 2024, 296, 154223. [Google Scholar] [CrossRef]
- Bantho, S.; Naidoo, Y.; Dewir, Y.H. The secretory scales of Combretum erythrophyllum (Combretaceae): Micromorphology, ultrastructure and histochemistry. S. Afr. J. Bot. 2020, 131, 104–117. [Google Scholar] [CrossRef]
- Stanilova, M.I.; Molle, E.D.; Yanev, S.G. Chapter 5 - Galanthamine production by Leucojum aestivum cultures in vitro. In The Alkaloids: Chemistry and Biology; Cordell, G.A., Ed.; Academic Press: New York, EEUU, 2010; Volume 68, pp. 167–270. [Google Scholar] [CrossRef]
- Bartmańska, A.; Tronina, T.; Popłoński, J.; Milczarek, M.; Filip-Psurska, B.; Wietrzyk, J. Highly cancer selective antiproliferative activity of natural prenylated flavonoids. Molecules. 2018, 23, 2922. [Google Scholar] [CrossRef]
- Wink, M. Ecological roles of alkaloids. In: Modern alkaloids: Structure, isolation, synthesis and biology, Fattorusso, E.; Taglialatela-Scafati, O., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA; Weinheim, Germany, 2008, pp. 3–52. [CrossRef]
- Kuntorini, E.M.; Nugroho, L.H. Structural development and bioactive content of red bulb plant (Eleutherine americana); a traditional medicines for local Kalimantan people. Biodiversitas. 2009, 11, 102–106. [Google Scholar] [CrossRef]
- Ferdausi, A.; Chang, X.; Hall, A.; Jones, M. Galanthamine production in tissue culture and metabolomic study on Amaryllidaceae alkaloids in Narcissus pseudonarcissus cv. Carlton. Ind. Crops Prod. 2020, 144, 112058. [Google Scholar] [CrossRef]
- Dewes-Neto, B.; Gomes-Copeland, K.K.P.; Silveira, D.; Gomes, S.M.; Craesmeyer, J.M.M.; de Castro Nizio, D.A.; Fagg, C.W. Influence of sucrose and activated charcoal on phytochemistry and vegetative growth in Zephyranthes irwiniana (Ravenna) Nic. García (Amaryllidaceae). Plants 2024, 13, 569. [Google Scholar] [CrossRef]
- Lubbe, A.; Gude, H.; Verpoorte, R.; Choi, Y.H. Seasonal accumulation of major alkaloids in organs of pharmaceutical crop Narcissus Carlton. Phytochemistry. 2013, 88, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Koirala, M.; Karimzadegan, V.; Liyanage, N.S., Mérindol, N.; Desgagné-Penix, I. Biotechnological approaches to optimize the production of Amaryllidaceae alkaloids. Biomolecules 2022, 12, 893. [CrossRef]
- Georgiev, V.; Ivanov, I.; Pavlov, A. Recent progress in Amaryllidaceae biotechnology. Molecules. 2020, 25, 4670. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S. Galantamine delivery for Alzheimer’s disease. In Sustainable Agriculture Reviews 43: Pharmaceutical Technology for Natural Products Delivery; Saneja, A., Panda, A.K., Lichtfouse, E., Eds.; Springer International Publishing: Berlin, Germany, 2020; Volume 1, pp. 131–159. [Google Scholar] [CrossRef]
- Petruczynik, A.; Misiurek, J.; Tuzimski, T.; Uszyński, R.; Szymczak, G.; Chernetskyy, M.; Waksmundzka-Hajnos, M. Comparison of different HPLC systems for analysis of galantamine and lycorine in various species of Amaryllidaceae family. J. Liq. Chromatogr. Relat. Technol. 2016, 39, 574–579. [Google Scholar] [CrossRef]
- Diamond, A.; Desgagné-Penix, I. Metabolic engineering for the production of plant isoquinoline alkaloids. Plant Biotechnol. J. 2016, 14, 1319–1328. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, Z.; Wang, S.; Li, J.; Li, Y.; Chen, W.K.; Wang, R. Fungal endophytes promote the accumulation of Amaryllidaceae alkaloids in Lycoris radiata. Environ. Microbiol. 2020, 22, 1421–1434. [Google Scholar] [CrossRef]
- Berkov, S.; Bastida, J.; Sidjimova, B.; Viladomat, F.; Codina, C. Alkaloid diversity in Galanthus elwesii and Galanthus nivalis. Chem. Biodivers. 2011, 8, 115–130. [Google Scholar] [CrossRef]
- Kaur, H.; Chahal, S.; Jha, P.; Lekhak, M.M.; Shekhawat, M.S.; Naidoo, D.; Arencibia, A.D.; Ochatt, S.J.; Kumar, V. Harnessing plant biotechnology-based strategies for in vitro galanthamine (GAL) biosynthesis: A potent drug against Alzheimer’s disease. Plant Cell Tiss. Organ Cult. 2022, 149, 81–103. [Google Scholar] [CrossRef]
- Sena, S.; Kaur, H.; Kumar, V. Lycorine as a lead molecule in the treatment of cancer and strategies for its biosynthesis using the in vitro culture technique. Phytochem. Rev. 2024, 1–28. [Google Scholar] [CrossRef]
- Yang, L. , Wen, K. S., Ruan, X., Zhao, Y. X., Wei, F., & Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules. 2018, 23, 762. [Google Scholar] [CrossRef]
- Roy, M.; Liang, L.; Xiao, X.; Feng, P.; Ye, M.; Liu, J. Lycorine: A prospective natural lead for anticancer drug discovery. Biomed. Pharmacother. 2018, 107, 615–624. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Qu, C.; Gao, O.; Hu, X.; Hong, X. Biological and pharmacological activities of Amaryllidaceae alkaloids. RSC Adv. 2015, 5, 16562–16574. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, P.; Sun, Y.; Sharopov, F.S.; Yang, Q.; Chen, F.; Wang, P; Liang, Z. Lycorine possesses notable anticancer potentials in on-small cell lung carcinoma cells via blocking Wnt/β-catenin signaling and epithelial-mesenchymal transition (EMT). Biochemical and biophysical research communications. 2018, 495, 911–921. [CrossRef]
- Wang, P.; Yuan, H.H.; Zhang, X.; Li, Y.P.; Shang, L.Q.; Yin, Z. Novel lycorine derivatives as anticancer agents: synthesis and in vitro biological evaluation. Molecules. 2014, 19, 2469–2480. [Google Scholar] [CrossRef] [PubMed]
- Johansen, D.A. Plant Microtechnique, MeGraw-Hill Book Company: New York, EEUU, 1940; pp. 1–523.
- Ruzin, S. Plant Microtechnique and Microscopy, Oxford University Press: Oxford, England, 1999; pp. 1–322.
- Tolivia, D.; Tolivia, J. Fasga: A new polychromatic method for simultaneous and differential staining of plant tissues. J. Microsc. 1987, 148, 113–117. [Google Scholar] [CrossRef]
- Demarco, D. Histochemical analysis of plant secretory structures. In Histochemistry of Single Molecules: Methods and Protocols, Methods in Molecular Biology, Pellicciari, C.; Biggiogera, M., Ed.; Humana: New York, EEUU, 2017; Volume 1560, pp. 313–330. [Google Scholar] [CrossRef]
- Brechú-Franco, A.E.; Laguna-Hernández, G.; De-La-Cruz-Chacón, I.; González-Esquinca, A.R. In situ histochemical localization of alkaloids and acetogenins in the endosperm and embryonic axis of Annona macroprophyllata Donn. Sm. seeds during germination. Eur. J. Histochem. 2016, 60, 2568. [Google Scholar] [CrossRef]
- Hernández, G.L.; Franco, A.B.; De-la-Cruz-Chacón, I.; González-Esquinca, A.R. Histochemical detection of acetogenins and storage molecules in the endosperm of Annona macroprophyllata Donn Sm. seeds. Eur. J. Histochem. 2015, 59, 2502. [Google Scholar] [CrossRef]
- Soto-Vásquez, M.R.; Rodríguez-Muñoz, C.A.; Tallini, L.R.; Bastida, J. Alkaloid composition and biological activities of the Amaryllidaceae species Ismene amancaes (Ker Gawl.). Herb. Plants. 2022, 11, 1906. [Google Scholar] [CrossRef]
- Cortes, N.; Sabogal-guaqueta, A.M.; Cardona-Gomez, G.P.; Osorio, E. Neuroprotection and improvement of the histopathological and behavioral impairments in a murine Alzheimer’s model treated with Zephyranthes carinata alkaloids. Biomed. Pharmacother. 2019, 110, 482–492. [Google Scholar] [CrossRef]
Figure 1.
P. lehmannii bulbs anatomy. A. Anatomic details of bulbs tissues; B. Amyloplast storage in the basal plate; C. Bud developing vascular tissue; D. Xylem vessels; E. Lysogenic cavities; F-J. Detection of alkaloids in the bud and scale leaves; F-G. Wagner's reagent; H. Lugol's reagent; I-J. Dragendorff's reagent. Arrow shows alkaloid storage: a = amyloplast, b = bud, bp = basal plate, lc = lysogenic cavities, r = root, s1 = scale leaf 1, s2 = scale leaf 2, s3 = scale leaf 3, t = tunic, vt = vascular tissue, xv = xylem vessels.
Figure 1.
P. lehmannii bulbs anatomy. A. Anatomic details of bulbs tissues; B. Amyloplast storage in the basal plate; C. Bud developing vascular tissue; D. Xylem vessels; E. Lysogenic cavities; F-J. Detection of alkaloids in the bud and scale leaves; F-G. Wagner's reagent; H. Lugol's reagent; I-J. Dragendorff's reagent. Arrow shows alkaloid storage: a = amyloplast, b = bud, bp = basal plate, lc = lysogenic cavities, r = root, s1 = scale leaf 1, s2 = scale leaf 2, s3 = scale leaf 3, t = tunic, vt = vascular tissue, xv = xylem vessels.
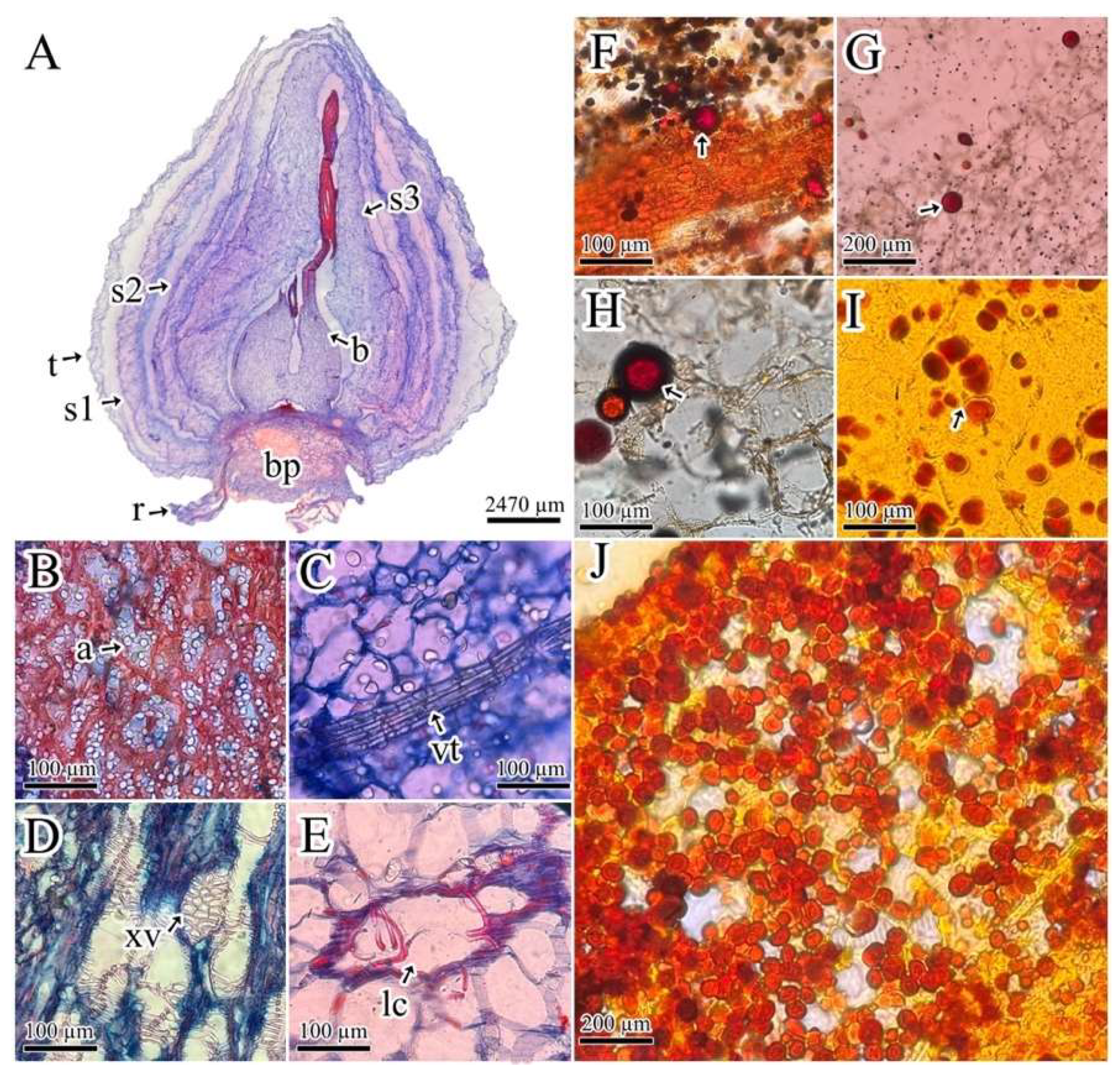
Figure 2.
Alkaloids identified in bulb and root from P. lehmannii.
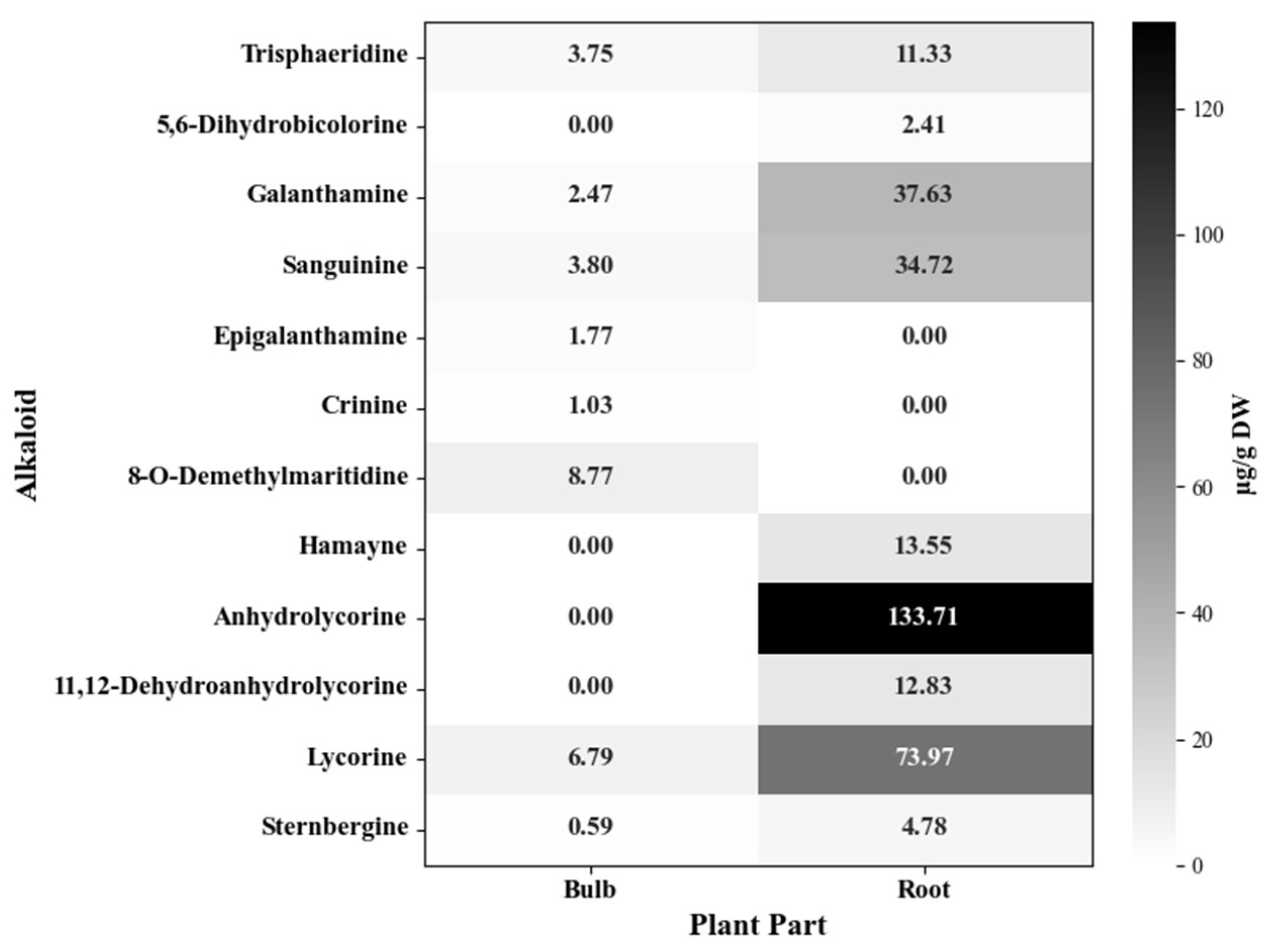
Figure 3.
Cell viability of alkaloid fractions of the bulb (A) and root (B) in gastric AGS and lung A549 cancer cells. Human keratinocytes HaCat were used as control cells. The results correspond to the mean ± SEM (n=3). ***P≤0.001 indicates statistically significant differences between control (DMSO 0. 1%) and treatments according to Dunnett's Multiple Comparison Test.
Figure 3.
Cell viability of alkaloid fractions of the bulb (A) and root (B) in gastric AGS and lung A549 cancer cells. Human keratinocytes HaCat were used as control cells. The results correspond to the mean ± SEM (n=3). ***P≤0.001 indicates statistically significant differences between control (DMSO 0. 1%) and treatments according to Dunnett's Multiple Comparison Test.
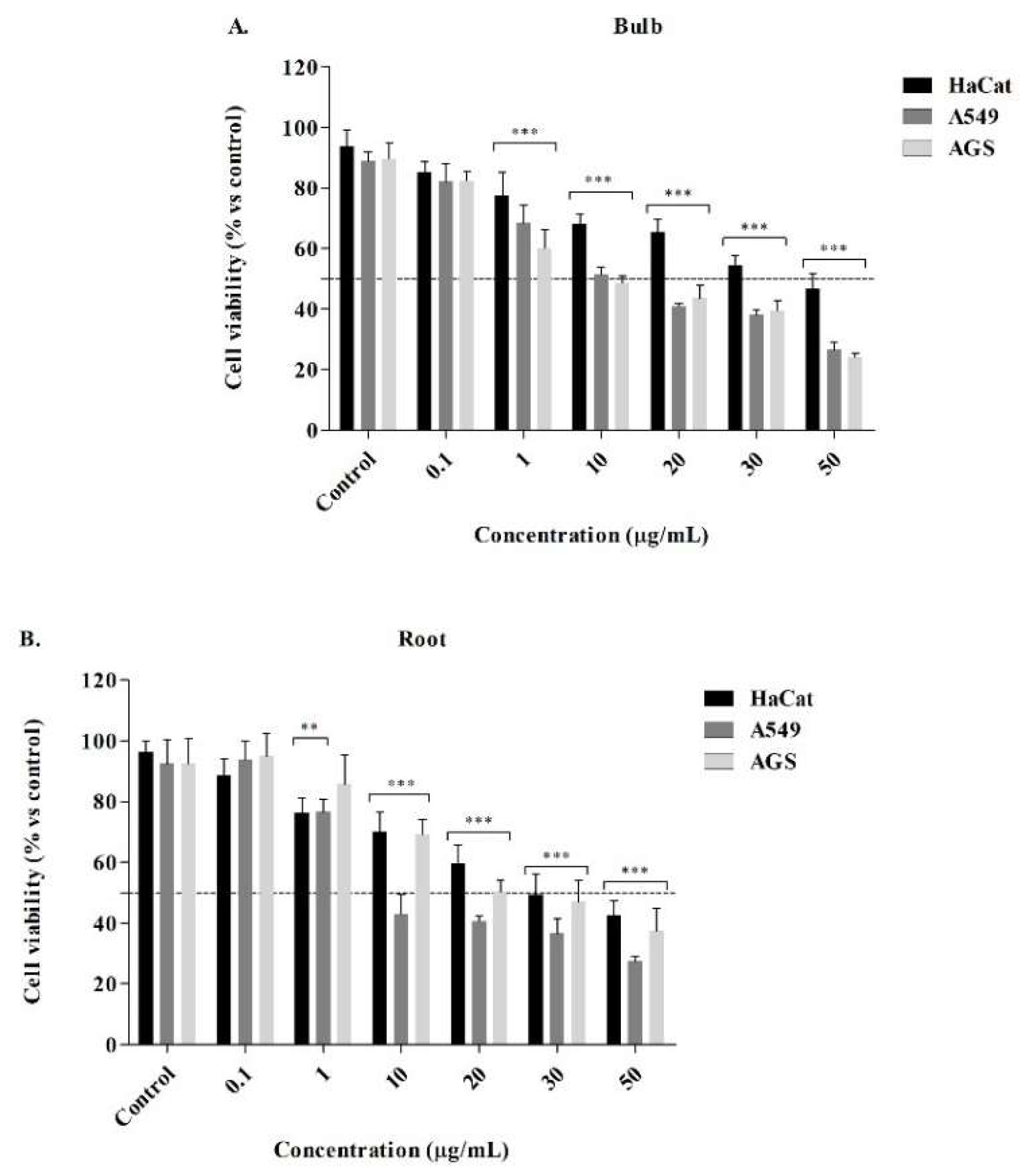

Table 1.
Cytotoxic potential of alkaloids P. lehmannii.
| IC50 (µg/mL) ± SD1 | |||||
|---|---|---|---|---|---|
| A5492 | SI4 | AGS2 | SI4 | HaCat3 | |
| Bulb | 11.76 ± 0.99 | 4.39 | 8.00 ± 1.35 | 6.45 | 51.59 ± 7.98 |
| Root | 2.59 ± 0.56 | 15.29 | 18.74 ± 1.99 | 2.11 | 39.60 ± 7.86 |
| Lycorine5 | 4.97 ± 0.89 | 7.92 | 4.07 ± 0.21 | 9.68 | 39.39 ± 0.94 |
| Doxorubicin5 | 5.52 ± 0.21 | 0.81 | 5.84 ± 0.65 | 0.77 | 4.48 ± 0.45 |
1 Results are the mean values standard deviations of three independent replications. 2 A549: lung cancer cell line and AGS: gastric cancer cell line. 3 HaCat: Human keratinocytes were used as control cells. 4 SI refers to the Selectivity Index (SI). 5 Positive controls.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



