
Submitted:
02 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Shigellosis is a pathological condition that affects the digestive system to possibly cause diarrhoea. Shigella species, which are responsible for this disease, are highly contagious and spread through contaminated food and water. The increasing development of resistance by Shigella species necessitate the urgent need to search for new therapies against shigellosis causing diarrhoea. The scientific validation of medicinal plants, such as Diospyros gilletii, which is used for the traditional treatment of diarrhoeal conditions is worthwhile. The present study aims to investigate the antibacterial activity of extracts from D. gilletii against selected Shigella species. Extracts from D. gilletii stem bark were prepared by maceration using various solvents. An-tibacterial activity of D. gilletii extracts was evaluated on Shigella dysenteriae, S. flexneri, S. boydii, and S. sonnei using a microdilution method, whereas cytotoxicity test was performed on Vero and Raw cells using resazurin-based colorimetric assay. Bacterial membrane permeability studies were evaluated using pro-pidium iodide (PI) and 1-N-phenylnaphthylamine (NPN) uptake assays, whereas inhibition and eradica-tion tests on bacterial biofilms were carried out by spectrophotometry. As a result, methanol, ethanol and hydroethanol extracts of D. gilletii inhibited the growth of S. boydii, S. flexneri and S. sonnei with MIC values ranging from 125 to 500 µg/mL, without toxicity to Vero and Raw cells. Time kill kinetics revealed bacteri-cidal orientation at 2MIC and 4MIC, and bacteriostatic outcome at 1/2MIC. The mechanistic basis of anti-bacterial action revealed that D. gilletii extracts inhibited and eradicated Shigella biofilms, and promoted the accumulation of NPN and PI within the inner and outer membrane of bacteria to increase membrane per-meability, thereby causing membrane damage. This novel contribution toward the antibacterial and mech-anisms of action of D. gilletii extracts against Shigella species substantiate the use of this plant in the tradi-tional treatment of infectious diarrhoea.

Keywords:
Diospyros gilletii
; Diarrhoea
; Anti-shigellosis activity
; Cytotoxicity
; Mechanisms of action
1. Introduction
Shigella spp. are gram negative and facultative anaerobic bacteria that can easily spread from one person to another and cause an enteric infection called shigellosis or bacillary dysentery [1]. Species of Shigella that are responsible for pathogenic diarrhoea include Shigella dysenteriae, Shigella flexneri, Shigella boydii and Shigella sonnei [2]. Infectious diarrhoea has been the cause of high morbidity and mortality in developing countries, especially in sub-Saharan Africa and South Asia [2,3]. Recent estimates pointed out about 188 million cases of Shigella infection per year with 1 million deaths. In fact, there are nearly 1.7 billion cases of childhood diarrhoeal disease every year, with approximately 525 000 under-five deaths [4]. Even children who survive diarrheal diseases face the likelihood of repeated infections and subsequent health issues over the rest of their lives. The 2015’s estimates point out 30,000,000 severe cases of diarrhea in Africa with 330,000 deaths [5]. In a study conducted by Njunda et al. [6], 4.5% of cases of Shigella infection was recorded among children under 15 years old after stool examination in the Buea Health District, Buea, and South West Region of Cameroon. It has also been reported that infantile diarrhea is among the main causes of death in children under 5 years old in Cameroon, together with malaria, measles and respiratory tract disease [7]. Humans are the main reservoir of Shigella bacteria ever since there is a well-established transmission from human to human by a fecal-oral route or ingestion of contaminated food and/or water [8]. When Shigella species enter into the digestive system, they are able to resist the acidity of the stomach to reach the small intestine and multiply. Next, Shigella migrates into the small intestine and produces enterotoxins and serotype toxin 1, which are responsible for watery and bloody diarrhoea. It takes only 12 hours to three days to observe the first clinical symptoms following contact with Shigella species [8]. Common manifestations of Shigella infection include vomiting, fever, tenesmus, headache, abdominal pain and bloody mucoid diarrhoeal [9,10]. Current treatments for infectious diarrhoea include the use of antibiotics, such as ciprofloxacin, ceftriaxone and azithromycin [11]. However, the development of resistance by Shigella species toward these antibiotics has resulted in their inefficiency [12]. In addition to drug resistance, the problem of toxicity has rendered these drugs useless. Thus, there is an urgent need to search for safe and effective anti-shigellosis drugs. Research on use of plants to treat infectious diseases has significantly contributed to the discovery of a number of anti-infective agents, including colchicine, emetine, quinine, and berberine, and so on. These essential medicines, which were powered by plants, have prompted many scientists to work on plants with the intention to develop safe therapies. For example, oregano oil encompasses the two most powerful anti-bacterial and antifungal compounds, namely carvacrol and thymol [13]. Numerous active antimicrobial principles (reserpine, opium, menthol and allicin), which were isolated respectively from plants, such as Vinca minor, Papaver somniferum, Mentha piperita, and Allium cepa, were found to inhibit the growth of a number of bacterial and fungal strains.
Based on these considerations, the search for safe antimicrobial agents might be inspired by active secondary metabolites from plants. One such plants include Diospyros gilletii, which was reported to contain norbergenin, 4-O-galloylnorbergenin, betulin, betulinic acid, lupeol, ursolic acid, corosolic acid, β-sitosterol and quercitol, inhibited Gram-positive (Bacillus subtilis DSMZ 704, Micrococcus luteus DSMZ 1605, Staphylococcus warneri DSMZ 20036) and Gram-negative (Escherichia coli DSMZ 1058 and Pseudomonas agarici DSMZ 11810) [14]. However, a detailed evaluation of the antibacterial activity of Diospyros gilletii stem bark against Shigella species, the pathogens responsible for infectious diarrhoea, is yet to be unveiled. Moreover, the use of Diospyros species in African folk medicine to treat various infectious diseases is undeniable [15,16].
Thus, the present study aims to investigate the antibacterial activity of Diospyros gilletii extracts against a number of Shigella species, including, Shigella flexneri, Shigella boydii, Shigella dysenteriae, and Shigella sonnei. The mechanistic basis of the antibacterial efficacy of D. gilletii extracts is also evaluated through biofilm inhibition and eradication, and membrane permeabilization assays.
2. Results
2.1. Yield of Extraction
The yields of extraction were found to be 5.6%, 7.61% and 8.27% for ethanol, methanol and hydroethanol extracts, respectively.
2.2. Antibacterial Activity
2.2.1. Minimum Inhibitory Concentrations (MICs)
Table 2 shows the MIC values corresponding to the lowest concentrations of the extracts for which there was no observed growth of Shigella spp. The in vitro incubation of Shigella species with different plant extracts has led to a decrease in the bacterial load as the minimum inhibitory concentrations varied from 125 to 1000 μg/ml. Methanol and hydroethanol extracts were found to be the most active extracts yielding MIC values of 125 and 250 μg/ml on Shigella boydii and Shigella sonnei, respectively. Shigella boydii (MIC<250 μg/ml) was found to be the most susceptible strain, whereas Shigella dysenteriae (MIC: 1000 μg/ml) was the most resistant strain. Ciprofloxacin, the standard antibiotic compound, presented MIC values varying from 0.010 to 0.039 μg/ml.
2.2.2. Time Kill Kinetics
The time kill-kinetics of methanol, ethanol and hydroethanol extracts were evaluated on the most susceptible strains, viz. S. flexneri NR 518, S. boydii NR 521, and S. sonnei NR 519. Figure 6, 7 and 8 shows the curves obtained by plotting optical densities (values that are proportional to the bacterial population) versus time of incubation of bacteria with methanol, ethanol and hydroethanol extracts, respectively. After two hours of bacterial incubation with the methanol extract, there was a decreasing trend of the curves, especially at MIC, 2MIC and 4MIC, irrespective of the Shigella strains considered (Figure 6). After 10 hours of incubation, at MIC, 2MIC and 4MIC, almost all the curves overlapped with the X axis (except for S. boydii), suggesting that the bacterial population was cleared, irrespective of the strain considered. This bacterial clearance was later confirmed as beyond 10 hours of incubation (10-24 hours), there was no resurgence of the curves, suggesting bactericidal orientation of the methanol extract at MIC, 2MIC and 4MIC. Treatment with ciprofloxacin showed the same trend, confirming the bactericidal trend of this drug vis-à-vis Shigella species. At a concentration less than the MIC (1/2 MIC), there was an increased inclination of the curve from 8 hours of incubation time onwards, suggesting a bacteriostatic orientation. By contrast, the curve obtained from the untreated group of bacteria showed an increased trend from time 0 to 24 hours, suggesting no inhibition and exponential growth of bacteria.
Figure 7 shows the growth kinetics of Shigella species after incubation with the ethanol extract of Diospyros gilleti. Irrespective of the concentration of extract considered (MIC, 2MIC and 4MIC), there was a decrease trend of the curves, suggesting inhibition of the bacterial population. After 10 hours of incubation at MIC, 2MIC and 4MIC, all the curves overlapped with the X axis, suggesting clearance of the bacterial population, irrespective of the strain considered. This bacterial clearance was later confirmed, since beyond 10 hours of incubation (10-24 hours), the bacteria did not resume growth, suggesting bactericidal orientation of the ethanol extract at MIC, 2MIC and 4MIC.
Figure 8 reveals the growth kinetics of Shigella species after incubation with the hydroethanol extract of Drospyros gilleti. At MIC, 2MIC and 4MIC, there was a decreased trend of the curves after 2 hours of incubation time, suggesting inhibition of the bacterial population. After 8 hours of incubation, all the curves overlapped with the X axis, suggesting clearance of the bacterial population. This bacterial clearance was later confirmed, since beyond 8 hours of incubation (8-24 hours), Shigella did not resume growth, suggesting bactericidal inclination of the hydroethanol extract at MIC, 2MIC and 4MIC.
2.2.3. Plausible Antibacterial Mechanisms of Action
Effect of Extracts on N-phenylnaphthylamine Uptake by Bacteria
The outer membrane permeability of the susceptible bacterial species (S. boydii, S. flexneri, and S. sonnei) was evaluated by measuring their 1-N-phenylnaphthylamine’s uptake upon treatment with different extracts of Diospyros gilleti stem bark. Figure 9 (A, B and C) reveals the effect of various concentrations of extracts (1/2MIC, MIC, 2MIC, and 4MIC) on the uptake of 1-N-phenylnaphthylamine in S. boydii, S. flexneri, and S. sonnei. At 2MIC and 4MIC, there was a significant increase in the relative fluorescence intensity from t0 to t90 minutes, suggesting higher intake of 1-N-phenylnaphthylamine by bacteria at these concentrations. A similar trend was observed when the bacterial strains were treated with ethylene diamine tetraacetic acid (EDTA), a compound that was used as a positive control. By contrast, the curves, which were obtained from the group of bacteria that did not receive any treatment with extracts remained unchanged throughout the experiment (from t0 to t90 minutes), inferring that untreated bacteria did not uptake 1-N-phenylnaphthylamine.
Propidium Iodide Uptake Assay
Following the graphs displayed in Figure 10 (A, B, and C), it can be seen that the fluorescence intensity, which was measured as a function of time, increased continuously for all treatments with extracts at 2MIC and 4MIC, vis-à-vis S. sonnei and S. boydii, suggesting propidium iodide (PI) uptake by these bacteria. Moreover, the curves obtained from treatments against S. flexneri remained unchanged throughout the experiment (0-70 minutes). Overall, methanol, ethanol and hydroethanol extracts triggered a rapid permeability of the bacterial membrane in S. sonnei and S. boydii, and propidium iodide fluorescence started increasing as from 10 minutes at 4 MIC and 2 MIC and thereafter, became constant while at 1 MIC and ½ MIC there was no increase in fluorescence on S. boydii and S. sonnei (Figure 10A, 10B and 10C). Normally, it is known that for extracts that work by propidium iodide uptake, the fluorescence intensity increases continuously. This shows that the antibacterial mode of action of the extracts is not achieved on S. flexneri through propidium iodide uptake. Nonetheless, this mode of action was verified on S. sonnei and S. boydii.
Inhibition and Eradication of Bacterial Biofilms
a. Inhibition of biofilm formation
According to Figure 11, the biofilm formation [by S. boydii (a) and S. sonnei (b)] was significantly inhibited (P≤0.05) by methanol, ethanol and hydroethanol extracts of D. gilletii stem bark (at MIC, 2MIC and 4MIC) after 24 h of incubation. The highest percentages of inhibition of the biofilm formation were observed with 4MIC. Moreover, ciprofloxacin, the positive control used, showed more than 100% inhibition of biofilms.
b. Eradication of biofilms
According to Figure 12, more than 50% of biofilms formed by S. bodyii (within 24 h of incubation) (Figure 12a) were eradicated by methanol, ethanol and hydroethanol extracts at MIC, 2MIC, 4MIC after 24 h of incubation. Moreover, the biofilms formed by S. sonnei (Figure 12b) were also eradicated (<50%) by all the extracts (except for methanol extract that showed 80% inhibition at 4MIC) at 4MIC, 2MIC and MIC.
2.2.4. Cytotoxicity Studies
Table 3 summarizes the 50% cytotoxic concentration of methanol (45.24 and 52.00 µg/ml), ethanol (54.52 and 65.30 µg/ml) and hydroethanol (39.94 and 47.15 µg/ml) extracts against Vero and Raw cells, respectively, vs dimethylsulfoxide (DMSO, a reference cytotoxic agent) (1.40 and 0.95 µg/ml). According to the National Cancer Institute classification, compounds with CC50 > 20 μg/ml are poorly cytotoxic or non-cytotoxic [17]. Compared to DMSO, which showed CC50 values less than 2 µg/ml, methanol, ethanol and hydroethanol extracts can be considered as non-cytotoxic extracts.
3. Discussion
Shigellosis is widely spread in underdeveloped or developing regions with poor hygiene and limited access to potable drinking water and has become a serious threat to public health. It is one of the most important causes of global childhood mortality and morbidity. Globally, there are about 165 million cases of Shigella infection with 1.1 million deaths annually [18]. Current treatments for Shigella infections include ciprofloxacin, ceftriaxone, and azithromycin [19]. However, Shigella species have become resistant to these medications [12], raising the urgent need to search for effective and safe antishigellosis agents. Plants have historically served as the most significant sources for new leads in drug discovery and development. Natural products are molecules produced by microbial agents, plants, and animals that have been demonstrated to be useful in treating microbial infections. Indeed, the World Health Organization estimated that nearly 75-80 % of the world population still depends on herbs and other traditional medicines for their primary health care needs. Among these plants, Diospyros species are widely used in African folk medicine for the treatment of various infectious diseases [15,16]. In the present study, extracts from the stem bark of Diospyros gilletii were evaluated for antibacterial activity against four species of Shigella, viz. S. boydii, S. sonnei, S. flexneri and S. dysentariae. Moreover, the antibacterial mechanism of action of the most active extracts was investigated. Solvent extraction of the stem bark of Diospyros gilletii yielded methanol, ethanol and hydroethanol extracts with yields of extraction of 5.6%, 7.61% and 8.27%, respectively. The hydroethanol, composed of two solvents i.e. ethanol and water might have induced a leakage of more secondary metabolites from the plant thereby increasing the yield of extraction (8.27%) [20]. The antibacterial test of the extracts revealed MIC values ranging from 125 to 1000 µg/ml, the most active extracts being the methanol and hydroethanol with MICs of 125 µg/ml when tested on S. boydii. According to previous reports [21,22,23], the degree of the antibacterial activity can be classified as very active (< 100 µg/ml), active (100 < MIC <512 µg/ml), moderately active (512 < MIC < 2048 µg/ml) and poorly active (MIC > 2048 µg/ml). Thus, the methanol extract was significantly active on S. flexneri (100 < MIC <512 µg/ml), whereas ethanol and hydro ethanol extracts were active (100 < MIC <512 µg/ml), on all the bacterial species. All the extracts showed poor activity against S. dysenteriae. The observed antibacterial activity might be due to the presence of a number of secondary metabolites [phenolic acids (betulinic acid, corosolic acid, ursolic acid and quercitol), flavonoids (norbergenin, 4-O-galloylnorbergenin), and the cholesterol derivative β-sitosterol] in Diospyros gilletii stem bark [14]. The different difference in the antibacterial action might be due to the diversity of the biochemical constituents and the genetic identity of bacteria, which potentially influenced the resistance in the tested bacteria [24]. Time kill kinetic studies revealed that the bacterial inhibition was proportional to the concentration of the extracts. In fact, the extracts were found to be bactericidal at 2MIC and 4MIC when tested against S. boydii, S. sonnei, and S. flexneri. To exert their activity within the cell, almost all antibiotics have to cross the outer membrane of bacteria either by increasing membrane permeability or its disruption. The measurement of the membrane permeability and disruption in bacteria might help in better understanding the mechanistic basis of the action [12]. The incubation of S. boydii, S. sonnei, and S. flexneri with various concentrations of extracts (4MIC, 2 MIC and MIC) promoted uptake of N-phenyl-naphthylamine and propidium iodide by these bacteria, leading to increase in the fluorescence [25]. Fluorescence of the NPN probe increases when this compound is incorporated into the hydrophobic core of the bacterial cell membrane as compared with the fluorescence of a non-permeating bacteria cell. The incubation of bacteria with D. gilletti extracts induced a significant accumulation of NPN in the bacterial membrane, which was further inhibited by free divalent cations, such as Mg2+ and Ca2+. These cations, which normally bind to negatively charged phosphate groups between lipopolysaccharide (LPS) molecules via ionic bridges, are crucial for the outer membrane integrity of Gram negative bacteria. Consequently, the chelation of these divalent ions by NPN would definitely disrupt the outer membrane’s integrity and permeabilize the bacteria [26]. Propidium iodide, a dye that labels bacteria with a compromised inner membrane, binds to nucleic acids and increases its fluorescent signal, and constitutes a strong indicator of inner membrane permeabilization. Methanol, ethanol and hydroethanol extracts promoted the PI and NPN uptake in S. boydii, and S. sonnei, inferring that the test extracts permeabilized inner and outer membrane of these bacteria [27,28]. Consequently, these extracts might have exerted antibacterial activity through bacterial membrane permeabilization. Extracts from Diospyros gilletii were further evaluated for antibiofilm formation by S. boydii, S. sonnei, and S. flexneri. The biofilm formation by bacteria has been intricately involved in the mechanism of bacterial resistance as the biofilm matrix can slow antibiotic penetration through the membrane. To avoid the development of microbial biofilms, inhibition of the bacterial adhesion, reduction of the initial bacterial growth and the eradication of formed biofilms are noteworthy. Methanol, ethanol and hydroethanol extracts from Diospyros gilletii inhibited the formation of biofilm with percentage of inhibition ranging from 25 % to 55 % on S. sonnei and from 13% to 90% on S. boydii. The observed anti-biofilm activity of the extracts might be attributed to the presence of a variety of secondary metabolites present in the plant [29]. Moreover, the extracts eradicated the biofilm formed by bacteria with percentages of inhibition varying from 8 % to 80 % on S. sonnei and from 48 % to 78 % on S. boydii. Anti-biofilm activity has been attributed to the potential of active compounds to interfere with biofilm development, destabilize the biofilm, promote detachment, and sensitize biofilm cells [30]. In S. boydii, ethanol and hydroethanol extracts inhibited and eradicated the biofilms. All the extracts eradicated biofilms formed by S. sonnei. For ethanol and hydroethanol extracts, the MBIC50 was found to be 4MIC, whereas the MBEC50 couldn’t be observed in any of the extracts. To evaluate the cytotoxicity profile of the active extracts, their median cytotoxic concentrations (CC50) were determined vis-à-vis human mammalian cells, such as Vero and Raw cells. The CC50 values ranged from 39.94±1.73 µg/ml to 54.525±2.73 µg/ml on Vero cells and from 47.15±10.38 µg/ml to 65.3±5.52 µg/ml on Raw cells. As the CC50 values of all the tested extracts were found to be more than 30 µg/ml, it can be speculated that none of the extracts was predicted to be toxic to the normal cells, according to the criteria of cytotoxicity defined by the American Nation Cancer Institute [31,32].
To the best of our knowledge, no report has demonstrated the antibacterial activity and mechanisms of action of methanol, ethanol and hydroethanol extracts from D. gilletii stem bark. The anti-Shigella activity of D. gilletii extracts, which is demonstrated in this study, substantiates the ethnopharmacological use of D. gilletti in the treatment of infectious diseases, such as infectious diarrhoea. However, antibacterial guided fractionation of D. gilletti extracts should be performed to identify the active anti-Shigella principles of this plant. In addition, in vivo anti-Shigella and pharmacokinetic studies, as well as detailed mechanisms of action are required for the successful utilization of D. gilletti in the treatment of infectious diarrhoea.
4. Material and Methods
4.1. Material
4.1.1. Plant Material
The stem bark of Diospyros gilletii De Wild was collected at Badounga, West Region of Cameroon in May 2022 and identified at the National Herbarium of Cameroon, where a voucher specimen of the species was deposited under number No 15418 HNC. The stem bark was then cut into small pieces, air-dried for two weeks and ground to obtain a powder.
4.1.2. Microbiological Material
The in vitro antibacterial activities activity of extracts from Diospyros gilletii stem bark was evaluated on three reference bacterial strains (Shigella flexneri NR 518, Shigella boydii NR 521 and Shigella sonnei NR 519), which were obtained from BEI resources and one clinical isolate, (Shigella dysenteriae), which was a kind gift from the Centre Pasteur of Cameroon. These bacterial strains were maintained in continuous culture in tubes on the Mueller Hinton Agar slope (4°C) at the Laboratory for Phytobiochemistry and Medicinal Plant Studies of the Faculty of Science of the University of Yaounde 1, Cameroon.
The in vitro cytotoxicity of the active extracts was evaluated on the mammalian African Green monkey Vero ATCC CRL 1586 and murine macrophage Raw cells 264.7 cell lines provided by the Centre Pasteur of Cameroon and from the Noguchi Memorial Institute for Medical Research, University of Ghana, respectively. The cells were maintained in a continuous culture in the laboratory and later used for the cytotoxicity test.
4.2. Methods
4.2.1. Antibacterial Activity
a. Preparation of crude extracts
The crude extracts were prepared by maceration of plant powder in three solvents, including ethanol, hydroethanol (30/70; v/v), and methanol. In brief, 10 g of dried powder were macerated in pure ethanol, methanol and hydroethanol (30/70; v/v) (100 ml) for 72 hrs at room temperature with daily shaking. The resulting solution was then filtered using a hydrophilic cotton, and the filtrate was further concentrated using a Buchi rotary evaporator at 55oC to yield the crude extracts. The extraction yield calculated using the following formula:
The crude extracts were kept in the refrigerator until further use.
b. Preparation of solutions
The stock solutions of the crude extracts were prepared at 100 mg/ml by dissolving 100 mg of each crude extract in 1ml of dimethlysulfoxide 100% in 1.5 ml Eppendorf tubes and stored at 4°C. Ciprofloxacin, which was used as the reference antibiotic, was prepared by dissolving 1 mg of drug in 1 ml of 0.5N HCL to yield a concentration of 1 mg/ml.
c. Preparation of bacterial inoculum
The different bacterial inoculum were prepared based on the 0.5 McFarland standard. Briefly, a colony from a 24 h culture on Müller Hinton agar (MHA) was removed using a platinum loop and introduce in a test tube containing 10 ml sterile physiological water (NaCl 0.9%), then calibrated to a 0.5 McFarland by turbidity comparison to obtain a 1.5×108UFC/ml.
d. Determination of minimum inhibitory concentrations
The determination of the MIC was done using the micro dilution method in liquid medium as described by the Clinical Laboratory Standard Institute (CLSI), M07-A9 protocol with slight modifications [33]. Briefly, 196 μl of Muller Hinton broth (MHB) were introduced in the first twelve wells of the plate and 100 μl in the rest wells of the microplate. Next, 4 μl of sterile solution containing different extracts concentrated at 100 mg/ml were introduced in the first twelve wells of column A, followed by a subsequent 5 two-fold serial dilution from line A to line F. Then, 100 μl of a bacterial suspension at 1x 106 CFU/ml was introduced in all the wells except for that of the sterility control. The negative control was composed of culture medium and inoculum only, while the positive control (concentration range: 2.5-0.15625 μg/ml) was made up of medium, inoculum and ciprofloxacin. Noteworthy, the final concentration of the inoculum in the wells was 5x105 CFU/ml. The microplates were subsequently incubated at 37o C for 24 h. At the end of the incubation period, 20 μl of a freshly prepared resazurin solution (0.15 mg/ml) was added in all the wells and further incubated for 30 min. The lowest concentration at which no color change from blue (resazurin) to pink (resorufin) was observed was considered as the minimum inhibitory concentration (MIC), corresponding to an absence of visible bacteria growth and was expressed in μg/mL. The test was done in triplicate and repeated twice. Subsequently, time kill kinetic assay was used to follow up the activity of plant extracts on the bacterial growth (bactericidal or bacteriostatic activity) over time.
4.2.2. Time Kill Kinetics
The time kill kinetic was studied using a modified protocol described by [34], notably the use of opacimetry based on the turbidity of the cell suspensions as a function of the load rather than colonies counting on agar. In short, the assay was performed in sterile 96-well micro plate at sub-inhibitory, inhibitory and supra-inhibitory extract concentrations. Briefly, the extracts were diluted in MHB medium as previously described in order to obtain ½ MIC, MIC, 2MIC and 4MIC. Ciprofloxacin (2.5 µg/ml) was used as a positive control. Then, 100 μl inoculum at 1×106 CFU/ml were introduced into each well to obtain a solution of 5 ×105 CFU/ml, except for the wells used for sterility control. The plates were then covered and incubated at 37oC for 24 h, during which the evolution of the bacterial load was followed by reading the optical densities at 630 nm of the preparations at different time intervals (0 , 1, 2, 4, 6, 8, 10, 12, and 24 hours ) using a TECAN infinite M200 microplate reader. The assay was done in triplicate and repeated twice. The bacterial growth curves were plotted using the optical densities against the incubation times. From these curves, we determined the minimum time at which the inhibitory effects started and the bactericidal and bacteriostatic effects of the extracts as well as the re-emergence time of the bacterial strains.
Next, plausible mechanisms of the antibacterial action of the most active extracts were investigated using membrane permeabilization test, and inhibitory/eradicative effect on bacterial biofilm formation.
4.2.3. Evaluation of Possible Mechanisms of Action
Membrane Permeabilization
The membrane permeabilization of the most promising extracts was evaluated using NPN (1-N- phenylnaphthylamine) and propidium iodide (PI) uptake assays.
a. 1-N-phenylnaphthylamine uptake assay
The outer membrane of gram-negative bacteria provides a formidable barrier (an asymmetric bilayer that contains mainly lipopolysaccharide (LPS) in the outer leaflet and phospholipids in the inner leaflet) that should be crossed by antimicrobial agents to exert their action. NPN (1-N-phenylnaphthylamine) is a neutral hydrophobic fluorescent probe that cannot permeate the intact outer membrane of the bacteria of which when the integrity of the bacteria outer membrane is destroyed, it will diffuse into the phospholipid bilayer and exhibit low fluorescence quantum yield in an aqueous environment and becomes strongly fluorescent in a glycerophospholipid or hydrophobic environment [27]. Briefly, the 1-N-phenylnaphthylamine (NPN) assay was performed to compare the effects of plant extract on the permeabilization of the outer membranes using a modified protocol of [26,27]. Briefly, in sterile 96-well black flat-bottomed micro plates bacterial cells were grown overnight, washed with normal saline (0.9% NaCl) and centrifuged at 10000 tr for 10 mins in eppendorfs tubes. The pellet was resuspended in normal saline, then adjusted to 0.5 Mcfarland in tubes containing glucose with phosphate buffer saline (50 mg of PBS in 50 ml of glucose). Next, 160 μl of cells were added to 20 μl NPN and 20 μl of extracts at various concentrations (4 MIC, 2 MIC, MIC, and ½ MIC). Positive control consisted of ethylene diamine tetraacetate (EDTA), NPN with bacteria suspension, whereas NPN was used as negative control. Sterility control was composed of NPN with normal saline. The micro plate was kept for 1h 30 mins in the dark and then NPN fluorescence was read at a 5 minutes interval using a TECAN infinite M200 micro plate reader at 350 nm and 420 nm excitation and emission wavelengths, respectively. The assay was performed in triplicate and repeated twice.
The percentage of NPN uptake was calculated using the following equation:
where:
Fobs = observed fluorescence at a given concentration of plant extract
F0= initial fluorescence of NPN with bacterial cells in the absence of plant extract
F100= fluorescence of NPN with bacteria cells upon addition of 2 μl of EDTA.
b. Propidium iodide uptake assay
The effect of antibacterial crude extracts on the bacterial membrane integrity was assayed using propidium iodide test. Propidium iodide (PI) is membrane-impermeable for intact cells and can only enter cells when the membrane permeability is augmented. Once in the cells, this fluorescent intercalating agent binds onto nucleic acids (DNA, RNA) of necrotic cells thereby discriminating between apoptotic and necrotic cells [28]. In this study, the PI uptake test was carried out according to a previously described protocol by Kwon et al. [27]. In sterile 96-well black flat-bottomed micro plates. Briefly, bacterial cells were grown overnight in sterile 96-well black flat-bottomed micro plates, washed with normal saline (0.9% NaCl) and then centrifuged at 10000 tr/10 mins in 1.5 mL Eppendorf tubes. The pellet was resuspended in normal saline then adjusted to 106 cells/mL from a 0.5Mcfarland for a final volume of 64000 μl in tubes containing 50 mg of PBS dissolved in 50 mL of glucose. Next, 20 μl of PI solution (1.3 mg of PI dissolved in 20 mL of PBS) was added to 160 μl of the bacterial suspension in a sterile 96-well black flat-bottomed micro plate and incubated in the dark for 30 minutes. Afterwards, 20 μl of various concentrations of plant extracts (4 MIC, 2 MIC, MIC, and ½ MIC) were added, while triton X was used as the positive control. Negative control was composed of propidium iodide and the bacterial suspension, while propidium iodide plus PBS was used as sterility control. Then, the microplate was incubated for 1 hour without light exposure and the optical densities were read using a TECAN infinite M200 microplate reader at 544 nm and 620 nm as excitation and emission wavelengths, respectively. The assay was performed in triplicate and repeated twice.
Antibiofilm Activity
Biofilms are structured microbial cells that occur as suspended aggregates attached to an inert or living surface to cause chronic infections [35]. Bacterial biofilms allow bacteria to resist and survive environmental pressure and enhance their resistance to antimicrobial agents [35].
a. Quantification of biofilm
Biofilm biomass measurement was performed by crystal violet staining according to a previously described protocol [36]. In brief, a single colony, which was isolated from an overnight bacteria culture on MHA was suspended in normal saline and adjusted to 1×106 CFU/ml in MHB supplemented with 2% glucose and 0.4% bovine bile. Next, 200 μl aliquot of the bacterial suspension was dispensed into a 96-well U shape sterile micro plate, whereas the negative control wells were filled with 200 μl MHB. The plates were then incubated for 24 hours at 37°C. At the end of the incubation period, the micro plate was gently emptied and 200 μl of pure ethanol were added to each well and incubated for 15 minutes in order to fix the cells in the wells. Next, the wells were then emptied and washed twice with sterile distilled water followed by an addition of 200 μl crystal violet 0.1% to stain the biofilm formed. The plates were further incubated at 37°C for 30 minutes, followed by removal of remaining (residual) crystal violet and three times washing of the wells using sterile distilled water. Finally, 200 μl of a 33% acetic acid solution were introduced into the wells and the absorbance was read at 590 nm using a TECAN (infinite M200) (Austria GmbH, Grödig Flachgau, Austria) plate reader.
b. Determination of the minimum inhibitory concentration (MBIC50) and minimum eradicative concentration (MBEC50) of biofilms
MBIC50 and MBEC50 were determined according to the micro dilution method in liquid medium. This assay was performed in triplicate in sterile 96-well micro plate at sub-inhibitory, inhibitory and supra-inhibitory extracts concentration. Briefly, the extracts were diluted in MHB medium supplemented with 2% glucose and 0.4% bovine bile salt as previously described to obtain 4 MIC, 2 MIC, MIC and ½ MIC. Next, a two-fold serial dilution of order 2 was carried out. Afterward, 100 μl of inoculum were added to all the wells and were then incubated for 24 hours at 37°C. The final bacterial load in each well was 1×106 CFU/ml. Ciprofloxacin (50 µg/mL) was used as positive control. After incubation, non-adherent or planktonic cells were discarded, and 200 μl of ethanol were added to each well in order to fix the cells in the wells and the micro plates were incubated for an additional 15 minutes. Then, the plates were gently washed (2 times) with distilled water and dried at room temperature. Next, each well was stained by adding 200 μl of 0.1% crystal violet and further incubated for 30 minutes, after which the stain was discarded and washed with distilled water so as to remove the excess stain. Finally, 200 μl of a 33% glacial acetic acid were introduced into the different wells and the absorbance was read at 590 nm using a TECAN (infinite M200) plate reader. The experiment was done in triplicate and repeated thrice. The MBEC50 was determined under the same conditions with the only difference being that the biofilms were first formed in the culture medium (MHB medium supplemented with 2% glucose and 0.4% bovine bile salt) for 24 h at 25°C. based on the strain. Next, the test samples were then prepared as described above and introduced into the corresponding wells of the microplates, already containing the formed biofilms. The microplates were further then re-incubated under the same conditions and then treated as elucidated above. Finally, 200 μl of a 33% glacial acetic acid were introduced into the different wells and the absorbance was read at 590 nm using a TECAN (infinite M200) plate reader. The experiment was done in triplicate and repeated thrice. The optical densities, which were obtained from the spectrophotometer were used to calculate the percentages of inhibition/eradication of biofilms using the following formula:
Where OD is the optical density.
MBIC50=minimal concentration that inhibited 50% biofilm formation, MBEC50 = minimal concentration that eradicated 50% of the biofilms formed.
Cytotoxicity studies of the most active extracts were carried out on Vero and Raw cells to evaluate their toxicity and selectivity.
4.2.4. Cytotoxicity Test
The extracts that inhibited the growth of the bacterial strains were evaluated against Vero ATCC CRL 1586 and Raw 264.7 cells using a colorimetric method based on the reduction of resazurin to resorufin [37].
In Vitro Culture of the Vero and Raw Cell Lines
The Vero and Raw cells were maintained in complete DMEM (Dulbecco’s Modified Eagle medium) (1% antibiotic and 10% FBS) and stored in a 25 cm3 cultured flask (T-Flask) under standard conditions of 5% CO2 at 37°C. The culture medium was renewed every 72 hrs. Once cell confluence (70-90%) was reached, the cells were detached by treatment with 1ml of trypsin-EDTA 0.05% for 5 min. After trypsin inactivation by adding 9 ml of complete DMEM, the detached cells were centrifuged at 1800 rpm for 5 min. The obtained pellet was then suspended in 1ml of the medium and the cell viability was evaluated using Trypan blue to calibrate the cell load on the Neubauer hemocytometer.
Determination of Median Cytotoxic Concentrations (CC50)
As already discussed, cytotoxicity of the most promising extracts was according to the method described by Bowling et al. [37]. The assay was performed in duplicate in sterile 96-well micro plates. Briefly, 100 μl of cells’ suspension containing 1×104 cells/well were introduced into all the wells of the plate and incubated for cell adhesion at 37°C, 5% CO2 for 18 hrs. Next, the cell culture medium was replaced with 90 μl of fresh medium followed by the addition of 10 μl of test samples at different concentrations (final concentrations of the extracts: 2000, 1000, 500, 250, 125, and 62.5 μg/ml) in all wells except for those that served as the positive (DMSO, 20 μM) and negative (culture medium without test sample) controls. The plates were then incubated at 37°C, 5% CO2 for 48 hours, after which 10 μl of a freshly prepared resazurin solution (0.15 mg/mL in PBS) were introduced into each well and incubated for 4 hours at 37°C in a 5% CO2. Next, the fluorescence of the obtained preparation was measured using TECAN microtiter plate reader (Infinite M200), at excitation and emission wavelengths of 530 and 590 nm, respectively. From the resulting values of optical densities, the percentage of cell viability was calculated with Microsoft excel software using the following equation:
where, At = Absorbance of test samples, Ac= Absorbance of negative control (cells without test samples).
A dose-response curve (cell viability versus concentration of the samples) was plotted using graph pad prism software, to determine the median cytotoxic concentration (CC50).
4.3. Statistical Analysis
The values were represented as the mean ± standard deviation. Statistical analysis was performed using the One-way Analysis of variance (ANOVA) using Graph Pad Prism 8.0.1 software. The percentages were calculated using Excel 2013 software. The differences between the means were compared by Fischer’s test with a 95% degree of confidence (p ≤ 0.05).
5. Conclusions and Perspectives
The present study aimed to evaluate the antibacterial activity and mechanisms of action of extracts from D. gilletti. Upon in vitro antibacterial test, extracts from D. gilletti afforded MIC values ranging 125 to 1000 µg/ml, with methanol extract being the most active extract because of its lowest MIC values (125 µg/ml). Potential antibacterial mechanistic studies of extracts from D. gilletti revealed that bactericidal orientation at 4MIC, and bacteriostatic tendency at 2MIC. Furthermore, the bioactive extracts contributed to the destabilization of the inner and outer membrane of the Shigella species through 1-N-phenylnaphthylamine and propidium iodide uptake by these membranes. In addition, D. gilletti extracts inhibited and eradicated biofilms formed by the shigellosis-causing microbes. D. gilletti extracts were also found non cytotoxic on Vero and Raw cells. These results validate the ethnomedicinal uses of D. gilletti in the traditional treatment of infectious diseases, such as diarrhoea. However, antibacterial guided fractionation of D. gilletti extracts should be performed to identify the active principles of this plant. Moreover, in vivo anti-shigellosis activity, pharmacokinetic and in depth mechanistic studies should be investigated for the successful utilization of D. gilletti in the treatment of diarrhoea caused by Shigella species.
Author Contributions
“Conceptualization, P.K.L., B.P.K. and F.F.B.; methodology, A.C.N.T., E.A.M.K., and B.-L.M.K.; software, E.A.M.K., B.-N.N.-D. and V.N.; validation, P.K.L., F.F.B., and B.P.K.; formal analysis, B.-L.M.K., L.M.N.N., Y.K.D.M., B.-N.N.-D and B.R.P.; investigation, A.C.N.T., B.-L.M.K., E.A.M.K., B.-N.N.-D and V.N; resources, P.K.L., F.F.B. and B.P.K.; data curation, A.C.N.T., B.-L.M.K., E.A.M.K., B.-N.N.-D, V.N., L.M.N.N., Y.K.D.M. and B.R.P.; writing—original draft preparation, A.C.N.T., E.A.M.K., B.-L.M.K., L.M.N.N., Y.K.D.M. and B.R.P.; writing—review and editing, P.K.L., B.P.K. and V.N.; visualization, P.K.L., B.P.K., V.N. and F.F.B; supervision, P.K.L. and F.F.B.; project administration, P.K.L. and F.F.B.; funding acquisition, P.K.L., B.P.K. and F.F.B. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research received external funding from the Yaounde–Bielefeld Bilateral Graduate School for Natural Products with Anti-parasite and Antibacterial Activity (YaBiNaPA) (grant number 57316173) and Seeding Labs’ Instrumental Access.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available from the corresponding author (s) upon reasonable request. The data are not publicly available due to the sensitive nature of the research supporting data.
Acknowledgments
Authors are thankful to the Cameroon National Herbarium (Yaounde, Cameroon) for their assistance with plant identification. The authors also acknowledge the ‘‘Centre Hospitalier Universitaire’’ of Yaounde–Cameroon and ‘‘Centre Pasteur’’ of Cameroon and for providing the bacterial strains species and the mammalian African Green monkey Vero ATCC CRL 1586. Thanks to the Noguchi Memorial Institute for Medical Research, University of Ghana, for providing the murine macrophage Raw cells 264.7 cell lines. This work was supported by and also received material and equipment support from the Yaoundé–Bielefeld Bilateral Graduate School for Natural Products with Anti-parasite and Antibacterial Activity (YaBiNaPA) and Seeding Labs’ Instrumental Access.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Al-Dahmoshi, H.; Al-Khafaji, N.; Al-Allak, M.; Salman, W.; Alabbasi, A. A review on shigellosis: Pathogenesis and antibiotic resistance. Drug Invent. Today 2020, 15(5) 793-798.
- Puzari, M.; Sharma, M.; Chetia, P. Emergence of antibiotic resistant Shigella species: A matter of concern. J. Infect. Public Health 2018, 11, 451-454. [CrossRef]
- Merzon, E.; Gutbir, Y.; Vinker, S.; Golan Cohen, A.; Horwitz, D.; Ashkenazi, S.; Sadaka, Y. Early childhood shigellosis and attention deficit hyperactivity disorder: A population-based cohort study with a prolonged follow-up. J. Atten. Disord. 2021, 25, 1791-1800. [CrossRef]
- The World Health Organization (WHO), 2023. Diarrhoeal disease. The Key Facts. https://www.who.int/news-room/fact-sheets/detail/diarrhoeal-disease, Accessed on 23rd December 2023.
- Reiner, R.C.Jr.; Graetz, N.; Casey, D.C.; Troeger, C.; Garcia, G.M.; Mosser, J.F.; Deshpande, A.; Swartz, S.J.; Ray, S.E.; Blacker, B.F.; Rao, P.C.; Osgood-Zimmerman, A.; Burstein, R.; Pigott, D.M.; Davis, I.M.; Letourneau, I.D.; Earl, L.; Ross, J.M.; Khalil, I.A.; Farag, T.H.; Brady, O.J.; Kraemer, M.U.G.; Smith, D.L.; Bhatt, S.; Weiss, D.J.; Gething, P.W.; Kassebaum, N.J.; Mokdad, A.H.; Murray, C.J.L.; Hay, S.I. Variation in childhood diarrheal morbidity and mortality in Africa, 2000-2015. N. Engl. J. Med. 2018, 379 (12) 1128-1138. [CrossRef]
- Njunda, A.L.; Assob, J.C.; Nsagha, D.S.; Kamga, H.L.; Awafong, M.P.; Weledji, E.P. Epidemiological, clinical features and susceptibility pattern of shigellosis in the buea health district, Cameroon. BMC Res. Notes 2012, 5, 54. [CrossRef]
- Ateudjieu, J.; Bita’a, L.; Guenou, E.; Chebe, A.; Chukuwchindun, B.; Goura, A.; Bisseck, A. Profile and antibiotic susceptibility pattern of bacterial pathogens associated with diarrheas in patients presenting at the Kousseri Regional Hospital Anne, Far North, Cameroon. Pan Afr. Med. J. 2018, 29 (170).
- Aslam, A.; Okafor, C.N. Shigella, in: StatPearls. StatPearls Publishing, Treasure Island (FL), 2022.
- Sheikh, A.F.; Moosavian, M.; Abdi, M.; Heidary, M.; Shahi, F.; Jomehzadeh, N.; Seyed-Mohammadi, S.; Saki, M.; Khoshnood, S. Prevalence and antimicrobial resistance of Shigella species isolated from diarrheal patients in Ahvaz, southwest Iran. Infect. Drug Resist. 2019, 12, 249-253. [CrossRef]
- Libby, T.E.; Delawalla, M.L.M.; Al-Shimari, F.; MacLennan, C.A.; Vannice, K.S.; Pavlinac, P.B. Consequences of Shigella infection in young children: a systematic review. Int. J. Infect. Dis. 2023, 129, 78-95. [CrossRef]
- 11. The World Health Organization (WHO), 2021: Global priority pathogens. Google Scholar [WWW Document]. URL https://scholar.google.com/scholar_lookup?author=World%20Health%20Organization&publication_year=2017&journal=WHO+global+priority+pathogens+list+of+antibiotic-resistant+bacteria (accessed 5.22.23).
- Ranjbar, R.; Farahani, A. Shigella: Antibiotic-resistance mechanisms and new horizons for treatment. Infect. Drug Resist. 2019, 12, 3137-3167. [CrossRef]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12(4), 564-582. [CrossRef]
- Tameye, N.S.J.; Akak, C.M.; Happi, G.M.; Frese, M.; Stammler, H.-G.; Neumann, B.; Lenta, B.N.; Sewald, N.; Nkengfack, A.E. Antioxidant norbergenin derivatives from the leaves of Diospyros gilletii De Wild (Ebenaceae). Phytochem. Lett. 2020, 36, 63-67. [CrossRef]
- Maroyi, A. Diospyros lycioides Desf.: Review of its botany, medicinal uses, pharmacological activities and phytochemistry. Asian Pac. J. Trop. Biomed. 2018, 8, 130. [CrossRef]
- Rauf, A.; Uddin, G.; Patel, S.; Khan, A.; Halim, S.; Bawazeer, S.; Ahmad, K.; Muhammad, N.; Mubarak, M. 2017. Diospyros, an under-utilized, multi-purpose plant genus: A review. Biomed. Pharmacother. 2017, 91, 714–730. [CrossRef]
- Anywar, G.U.; Kakudidi, E.; Oryem-Origa, H.; Schubert, A.; Jassoy, C. Cytotoxicity of medicinal plant species used by traditional healers in treating people suffering from HIV/AIDS in Uganda. Front. Toxicol. 2022 May 2;4:832780. [CrossRef]
- Kotloff, K.L.; Riddle, M.S.; Platts-Mills, J.A.; Pavlinac, P.; Zaidi, A.K.M. Shigellosis. The Lancet 2018, 391, 801-812.
- Lefèvre, S.; Njamkepo, E.; Feldman, S.; Ruckly, C.; Carle, I.; Lejay-Collin, M.; Fabre, L.; Yassine, I.; Frézal, L.; Pardos de la Gandara, M.; Fontanet, A.; Weill, F.X. Rapid emergence of extensively drug-resistant Shigella sonnei in France. Nat. Commun. 2023, 14 (1) 462. [CrossRef]
- Nur Syukriah, A.R.; Liza, M.S.; Harisun, Y.; Fadzillah, A.A.M. Effect of solvent extraction on antioxidant and antibacterial activities from Quercus infectoria (Manjakani). Int. Food Res. J. 2014, 21(3) 1067-1073.
- Tamokou, J.D.D.; Mbaveng, A.; Kuete, V. 2017. Antimicrobial activities of African medicinal spices and vegetables, in: Medicinal Spices and Vegetables from Africa: Therapeutic Potential Against Metabolic, Inflammatory, Infectious and Systemic Diseases. pp. 207-237. [CrossRef]
- Franklin Loic, T.T.; Boniface, P.K.; Vincent, N.; Zuriatou Y.T.; Victorine Lorette, Y.; Julius, N.N.; Paul, K.L.; Fabrice, F.B. Biological synthesis and characterization of silver-doped nanocomposites: Antibacterial and mechanistic studies. Drugs Drug Candidates 2024, 3, 13-32.
- Brice Rostan P.; Boniface, P.K.; Eutrophe Le Doux K.; Vincent N.; Yanick Kevin M.D.; Paul, K.L.; Fabrice, F.B. Extracts from Cardiospermum grandiflorum and Blighia welwitschii (Sapindaceae) reveal antibacterial activity against Shigella species. S. Afr. J. Bot. 2024, 164, 419-428.
- Van Bambeke, F.; Mingeot-Leclercq, M.-P.; Struelens, M.J.; Tulkens, P.M. The bacterial envelope as a target for novel anti-MRSA antibiotics. Trends Pharmacol. Sci. 2008, 29, 124-134. [CrossRef]
- Wang, Q.; Kim, H.; Halvorsen, T.M.; Buie, C.R. Leveraging microfluidic dielectrophoresis to distinguish compositional variations of lipopolysaccharide in E. coli (preprint). Front. Bioeng. Biotechnol. 2023, 11, 991784. [CrossRef]
- Loh, B.; Grant, C.; Hancock, R.E. Use of the fluorescent probe 1-N-phenylnaphthylamine to study the interactions of aminoglycoside antibiotics with the outer membrane of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1984, 26, 546-551. [CrossRef]
- Kwon, J.Y.; Kim, M.K.; Mereuta, L.; Seo, C.H.; Luchian, T.; Park, Y. Mechanism of action of antimicrobial peptide P5 truncations against Pseudomonas aeruginosa and Staphylococcus aureus. AMB Express. 2019, 9, 122. [CrossRef]
- Van Moll, L.; De Smet, J.; Paas, A.; Tegtmeier, D.; Vilcinskas, A.; Cos, P.; Van Campenhout, L. In vitro evaluation of antimicrobial peptides from the black soldier fly (Hermetia Illucens) against a selection of human pathogens. Microbiol. Spectr. 2022, 10, e01664-21. [CrossRef]
- Qu, Y.; Locock, K.; Verma-Gaur, J.; Hay, I.D.; Meagher, L.; Traven, A. Searching for new strategies against polymicrobial biofilm infections: guanylated polymethacrylates kill mixed fungal/bacterial biofilms. J. Antimicrob. Chemother. 2016, 71(2) 413-421. [CrossRef]
- Wolfmeier H, Pletzer D, Mansour SC, Hancock REW. New perspectives in biofilm eradication. ACS Infect. Dis. 2018, 4(2), 93-106. [CrossRef]
- Ogbole, O.O.; Segun, P.A.; Adeniji, A.J. In vitro cytotoxic activity of medicinal plants from Nigeria ethnomedicine on Rhabdomyosarcoma cancer cell line and HPLC analysis of active extracts. BMC Complement. Altern. Med. 2017, 17, 494. [CrossRef]
- Serrano-Vega, R.; Pérez-Gutiérrez, S.; Alarcón-Aguilar, F.; Almanza-Pérez, J.; Pérez-González, C.; González-Chávez, M.M. Phytochemical composition, anti-inflammatory and cytotoxic activities of chloroform extract of Senna crotalarioides Kunth. Am. J. Plant Sci. 2021, 12, 887-900. [CrossRef]
- CLSI, 2012. CLSI Publishes 2012 Antimicrobial Susceptibility Testing Standards - Medical Design and Outsourcing. https://www.medicaldesignandoutsourcing.com/clsi-publishes-2012-antimicrobial-susceptibility-testing-standards/ (accessed 6.7.23).
- Klepser, M.E.; Ernst, E.J.; Lewis, R.E.; Ernst, M.E.; Pfaller, M.A. Influence of test conditions on antifungal time-kill curve results: proposal for standardized methods. Antimicrob. Agents Chemother. 1998, 42, 1207-121. [CrossRef]
- Liu, X.; Yao, H.; Zhao, X.; Ge, C. Biofilm formation and control of foodborne pathogenic bacteria. Molecules 2023, 28, 243. [CrossRef]
- Kalia, M.; Singh, D.; Sharma, D.; Narvi, S.; Agarwal, V. Senna alexandriana mill as a potential inhibitor for quorum sensing-controlled virulence factors and biofilm formation in Pseudomonas aeruginosa PAO1. Phcog. Mag. 2020, 16, 80. [CrossRef]
- Bowling, T.; Mercer, L.; Don, R.; Jacobs, R.; Nare, B. Application of a resazurin-based high-throughput screening assay for the identification and progression of new treatments for human African trypanosomiasis. Int. J. Parasitol. Drugs Drug Resist. 2012, 2, 262-27. [CrossRef]
Figure 6.
Effects of DGs Drospyros gilleti methanolic extracts on the growth kinetics of S. boydii, S. flexneri, and S. sonnei. DGsM: Methanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
Figure 6.
Effects of DGs Drospyros gilleti methanolic extracts on the growth kinetics of S. boydii, S. flexneri, and S. sonnei. DGsM: Methanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
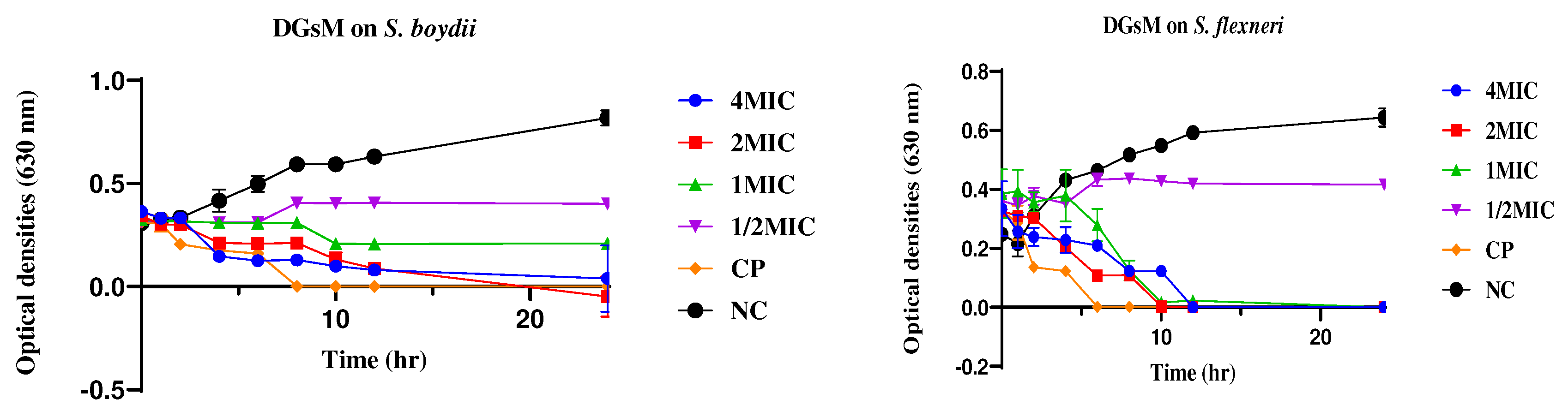
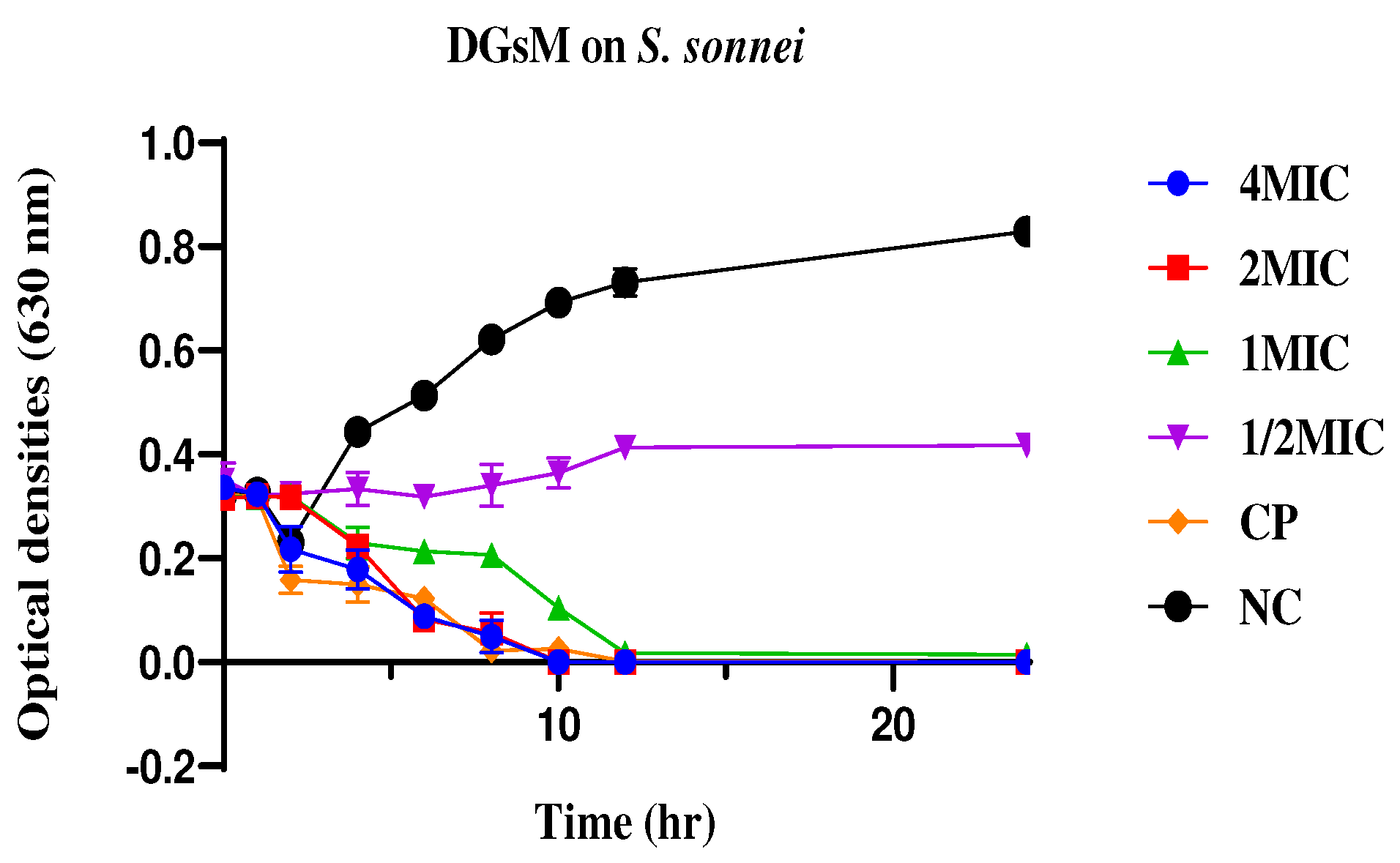
Figure 7.
Effects of Drospyros gilleti DGs ethanolic extracts on the growth kinetics of S. boydii, S. flexneri, and S. sonnei. DGsE: Ethanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
Figure 7.
Effects of Drospyros gilleti DGs ethanolic extracts on the growth kinetics of S. boydii, S. flexneri, and S. sonnei. DGsE: Ethanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
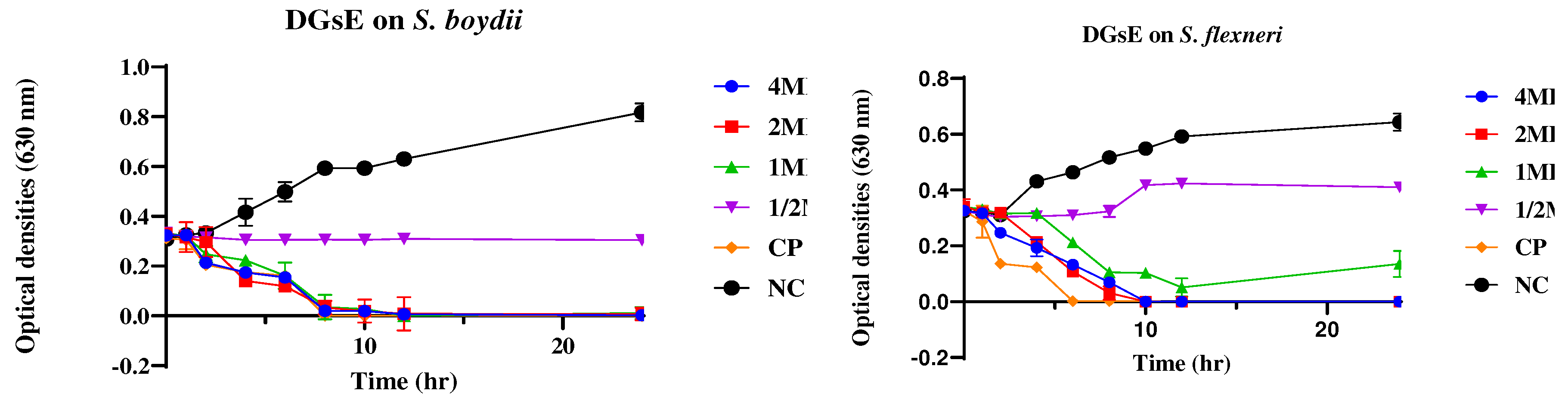

Figure 8.
Effects of the hydroethanol extract of Drospyros gilletii on the growth kinetics of S. boydii, S. flexneri and S. sonnei. DGsHE: Hydrothanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
Figure 8.
Effects of the hydroethanol extract of Drospyros gilletii on the growth kinetics of S. boydii, S. flexneri and S. sonnei. DGsHE: Hydrothanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
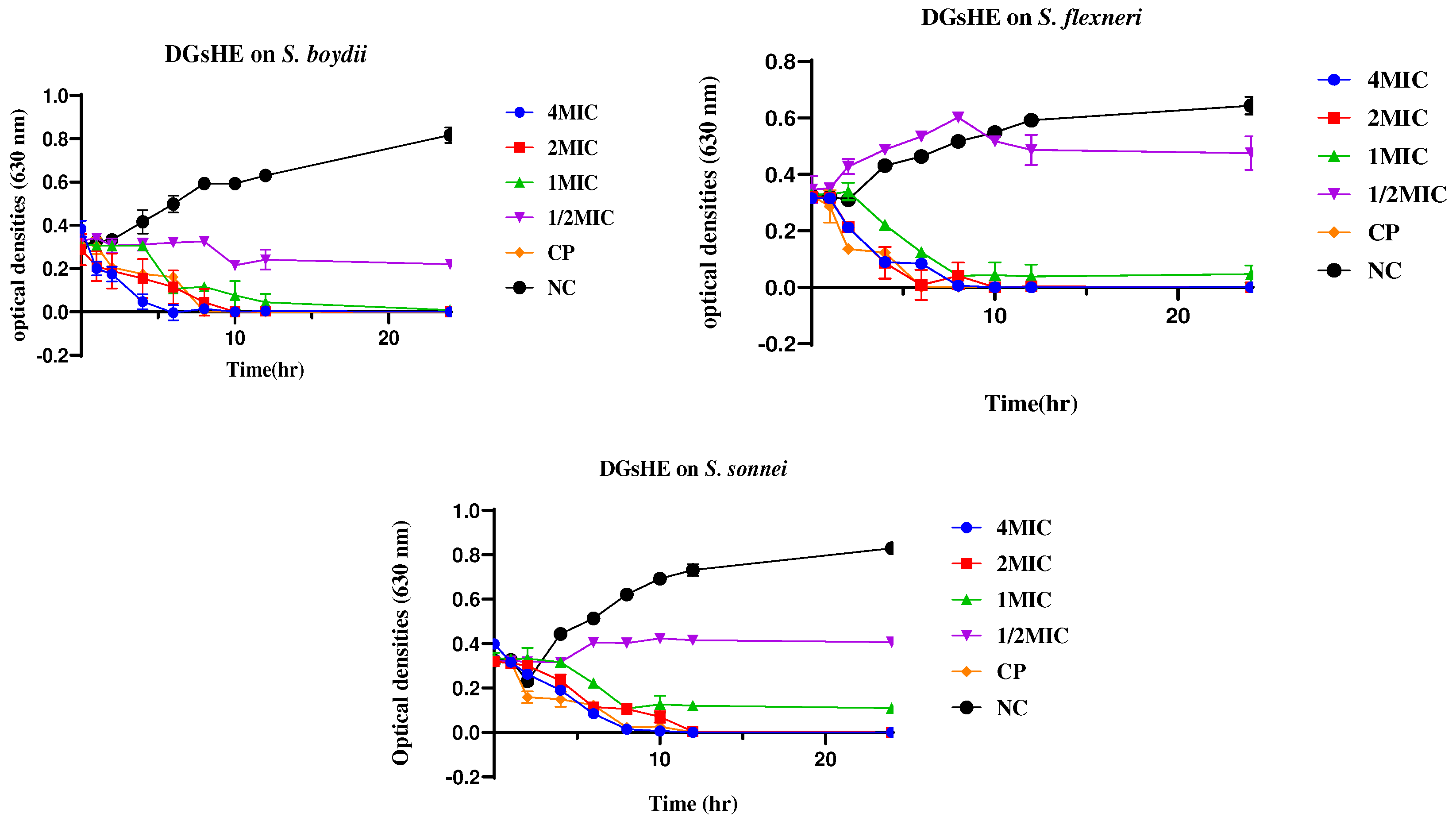
Figure 9.
Effect of various concentrations of methanol (A), ethanol (B) and hydroethanol (C), and extracts on the uptake of N-phenylnaphthylamine by S. boydii, S. flexneri, and S. sonnei. DGsE: Ethanol extract of Drospyros gilleti; DGsHE: Hydroethanol extract of Drospyros gilleti; DGsM: Methanol extract of Drospyros gilleti; EDTA : ethylene diamine tetraacetic acid ; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
Figure 9.
Effect of various concentrations of methanol (A), ethanol (B) and hydroethanol (C), and extracts on the uptake of N-phenylnaphthylamine by S. boydii, S. flexneri, and S. sonnei. DGsE: Ethanol extract of Drospyros gilleti; DGsHE: Hydroethanol extract of Drospyros gilleti; DGsM: Methanol extract of Drospyros gilleti; EDTA : ethylene diamine tetraacetic acid ; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
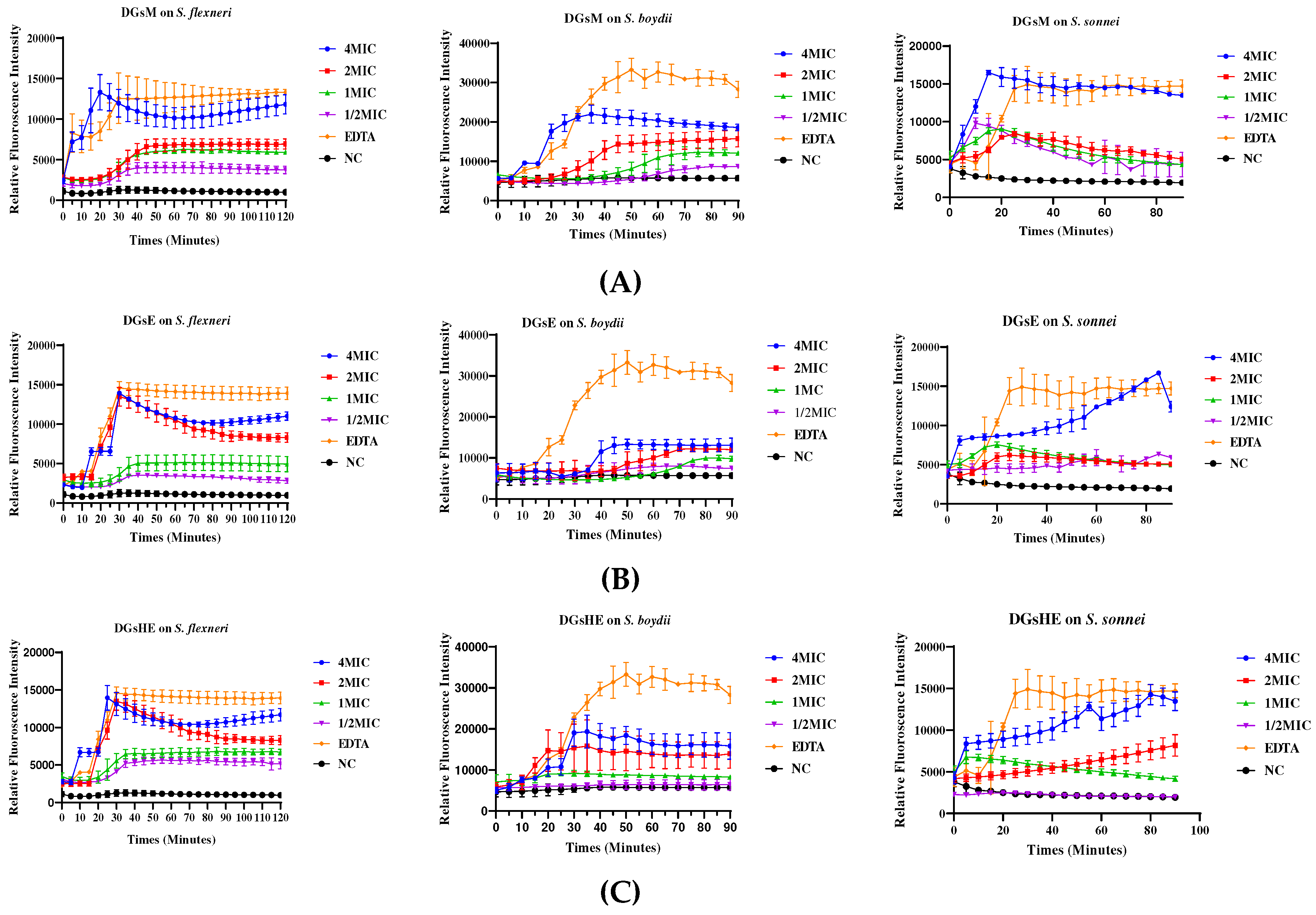
Figure 10.
Effect of various concentrations of methanol (A), ethanol (B) and hydroethanol (C), and extracts on the uptake of propidium iiodide by S. boydii, S. flexneri, and S. sonnei. DGsE: Ethanol extract of Drospyros gilleti; DGsHE: Hydroethanol extract of Drospyros gilleti; DGsM: Methanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
Figure 10.
Effect of various concentrations of methanol (A), ethanol (B) and hydroethanol (C), and extracts on the uptake of propidium iiodide by S. boydii, S. flexneri, and S. sonnei. DGsE: Ethanol extract of Drospyros gilleti; DGsHE: Hydroethanol extract of Drospyros gilleti; DGsM: Methanol extract of Drospyros gilleti; MIC : Minimum inhibitory concentration ; NC : Negative control ; CP : ciprofloxacin ; S. boydii: Shigella boydii; S. flexneri: Shigella flexneri; S. sonnei: Shigella sonnei.
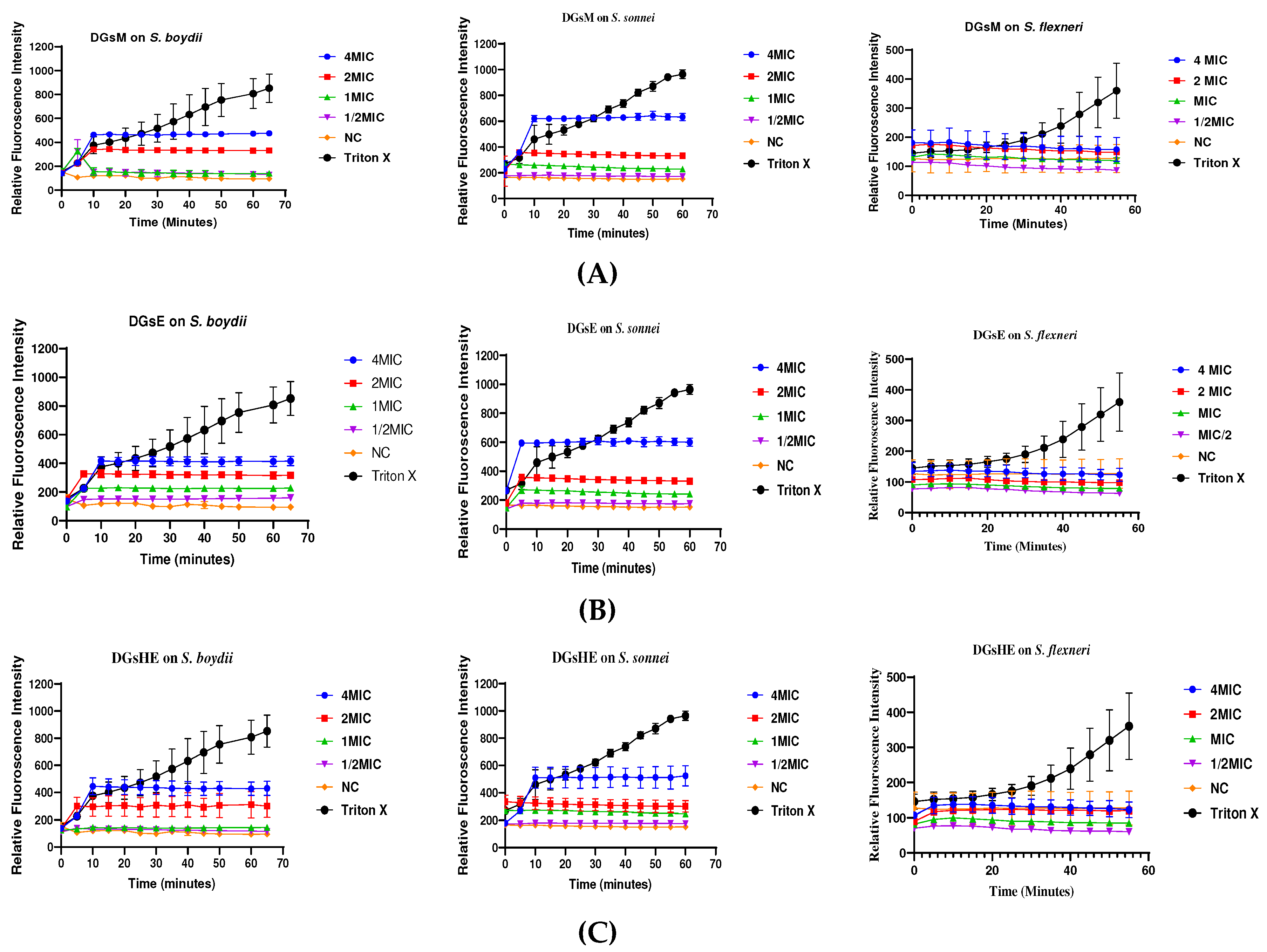
Figure 11.
Inhibition of biofilms from S. bodyii (a) and S. sonnei (b) by methanol, ethanol and hydroethanol extracts. Extract 1: Methanol extract; Extract 2: Ethanol extract; Extract 3: Hydroethanol extract; MIC: Minimum inhibitory concentration. aP ≤ 0.05, bP ≤ 0.01, cP ≤ 0.001; Values are significantly different, compared to the value of negative control (no biofilm inhibition).
Figure 11.
Inhibition of biofilms from S. bodyii (a) and S. sonnei (b) by methanol, ethanol and hydroethanol extracts. Extract 1: Methanol extract; Extract 2: Ethanol extract; Extract 3: Hydroethanol extract; MIC: Minimum inhibitory concentration. aP ≤ 0.05, bP ≤ 0.01, cP ≤ 0.001; Values are significantly different, compared to the value of negative control (no biofilm inhibition).
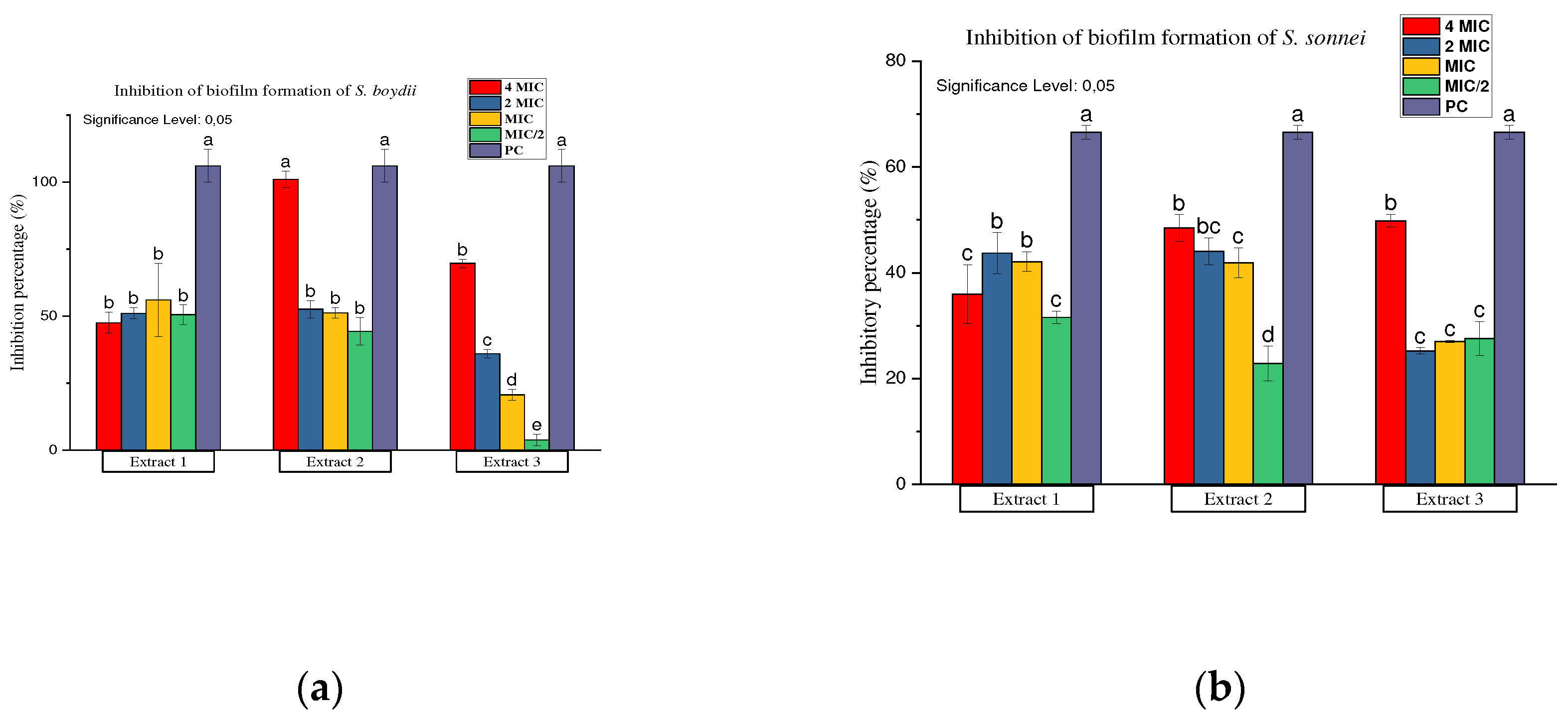
Figure 12.
Eradication of biofilms formed by S. bodyii and S. sonnei using methanol, ethanol and hydroethanol extracts. Extract 1: Methanol extract; Extract 2: Ethanol extract; Extract 3: Hydroethanol extract; MIC: minimum inhibitory concentration. aP ≤ 0.05, bP ≤ 0.01, cP ≤ 0.001; Values are significantly different, compared to the value of negative control (no biofilm eradication).
Figure 12.
Eradication of biofilms formed by S. bodyii and S. sonnei using methanol, ethanol and hydroethanol extracts. Extract 1: Methanol extract; Extract 2: Ethanol extract; Extract 3: Hydroethanol extract; MIC: minimum inhibitory concentration. aP ≤ 0.05, bP ≤ 0.01, cP ≤ 0.001; Values are significantly different, compared to the value of negative control (no biofilm eradication).
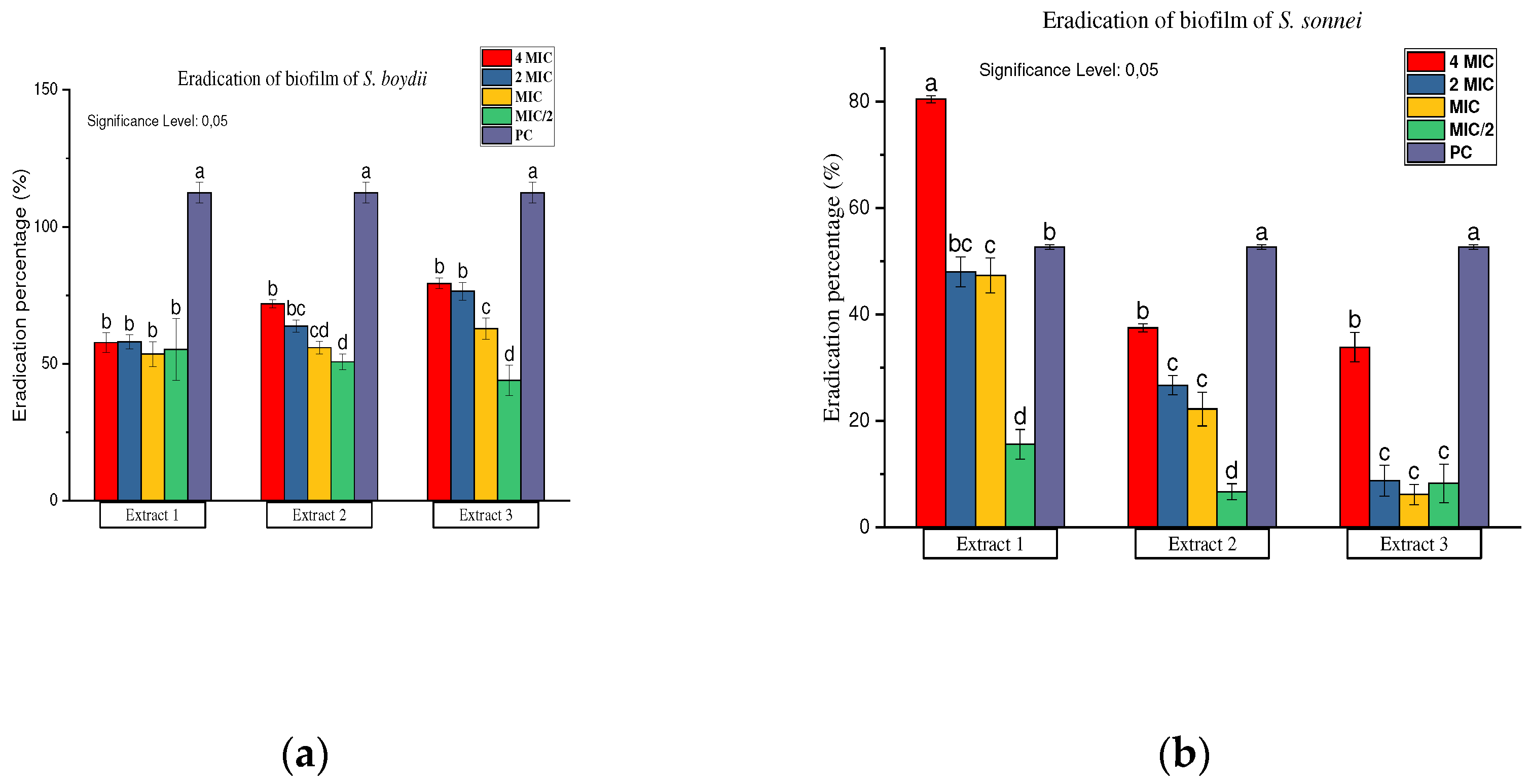

Table 2.
Minimum inhibitory concentration of extracts on Shigella spp.
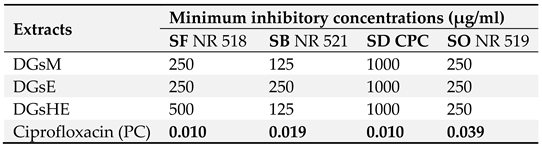 |
CPC : Centre Pasteur of Cameroon ; DGsE : Ethanol extract of Drospyros gilleti ; DGsHE : Hydroethanol extract of Drospyros gilleti; DGsM: Methanol extract of Drospyros gilleti; PC : Positive control ; SB: Shigella boydii; SD: Shigella dysenteriae ; SF: Shigella flexneri; SO: Shigella sonnei.
Table 3.
Median cytotoxic concentrations of the extracts.
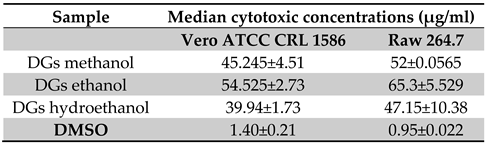 |
DGs Methanol: Methanol extract of Diospyros gilleti, DGs Ethanol: Ethanol extract of Diospyros gilleti; DGs Hydroethanol: Hydroethanol extract of Diospyros gilleti; DMSO: Dimethylsulfoxide.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



